The potassium channel subunit KV1.8 (Kcna10) is essential for the distinctive outwardly rectifying conductances of type I and II vestibular hair cells
- Department of Neurobiology, University of Chicago, United States
- Department of Anatomy and Cell Biology, University of Illinois at Chicago, United States
Figures
Figure 1 with 1 supplement
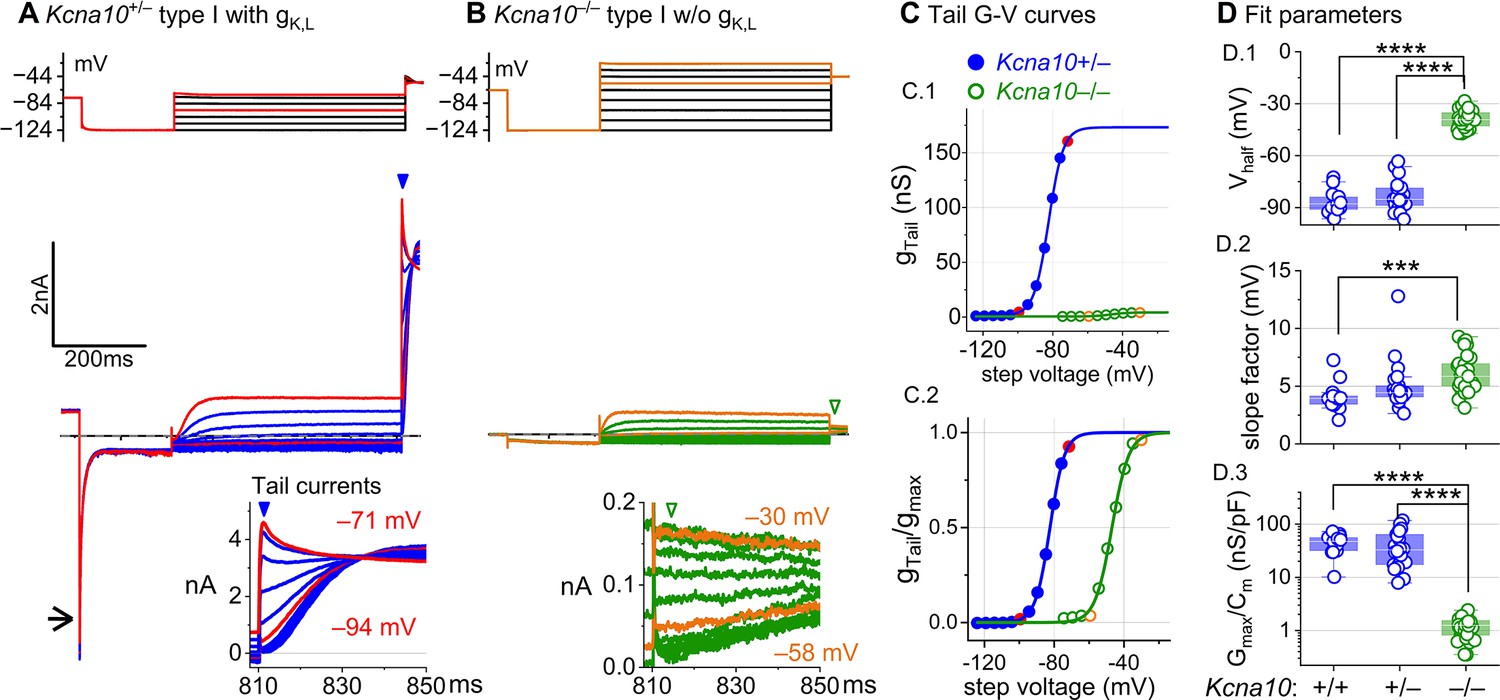
Kcna10–/– type I hair cells (HCs) lacked gK,L, the dominant conductance in mature Kcna10+/+,++/–– type I HCs.
Representative voltage-evoked currents in (A) a P22 Kcna10+/– type I HC and (B) a P29 Kcna10–/– type I HC. (A) Arrow, transient inward current that is a hallmark of gK,L. Arrowheads, tail currents, magnified in insets. For steps positive to the midpoint voltage, tail currents are very large. As a result, K+ accumulation in the calyceal cleft reduces driving force on K+, causing currents to decay rapidly, as seen in A (Lim et al., 2011). Note that the voltage protocol (top) in B extends to more positive voltages. (C) Activation (G–V) curves from tail currents in A and B; symbols, data; curves, Boltzmann fits (Equation 1). (D) Fit parameters from mice >P12 show big effect of Kcna10–/– and no difference between Kcna10+/– and Kcna10+/+. (D.1), Tukey’s test: +/+ vs –/–, p<1E-9; +/– vs –/–, p<1E-9. (D.2), Tukey’s test: +/+ vs –/–, p=9.4E-4. (D.3), Tukey’s test: +/+ vs –/–, p<1E-9; +/– vs –/–, p<1E-9. Asterisks: ***p < 0.001; and ****p < 0.0001. Line, median; Box, interquartile range; Whiskers, outliers. See Table 1 for statistics.
Figure 1—figure supplement 1
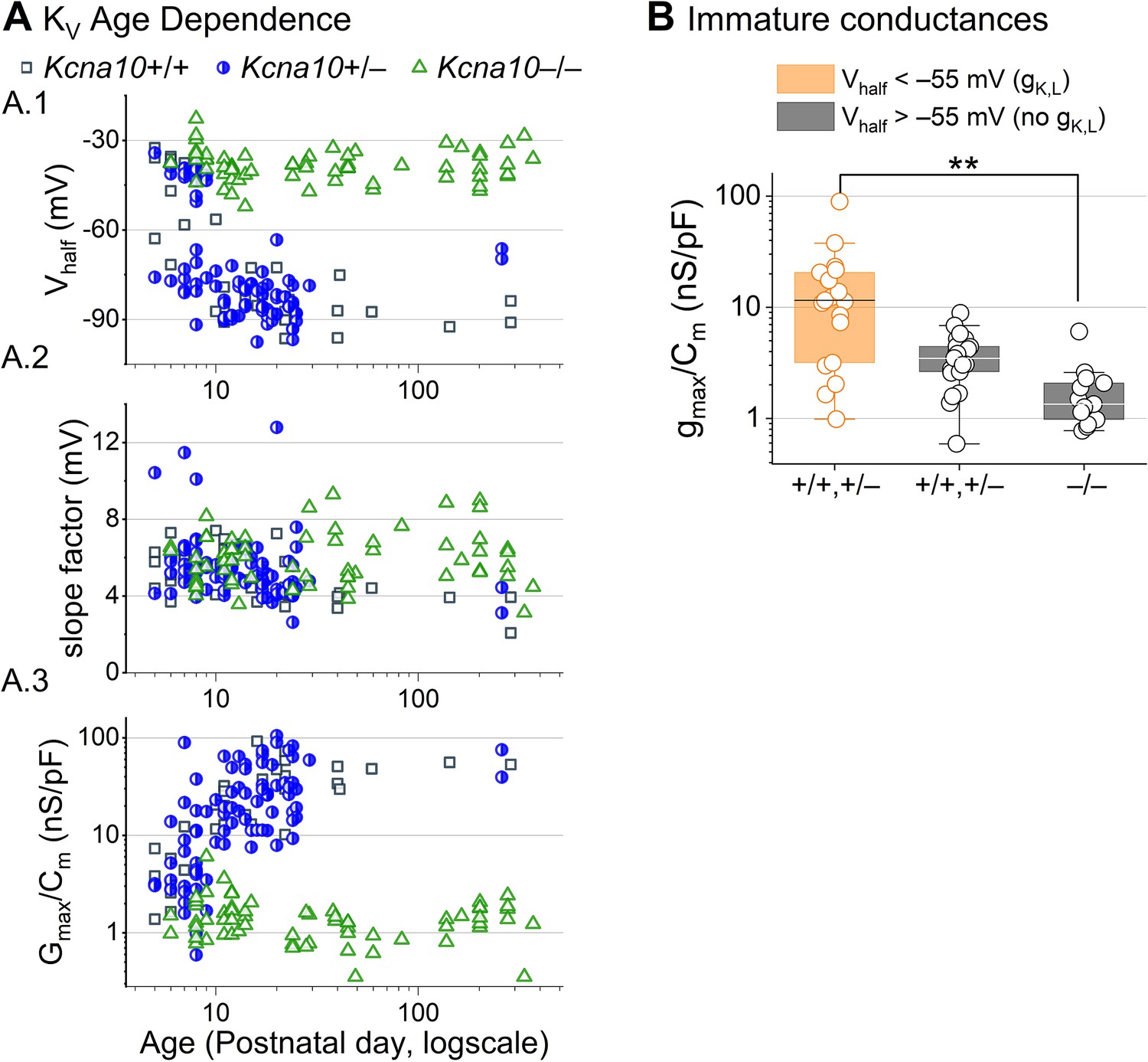
Developmental changes in type I hair cell (HC) KV conductances.
(A) Parameters from Boltzmann fits of tail G–V relations for type I HCs plotted against age. (B) Conductance density is similar in young (P5–P10) type I HCs that lack gK,L. gK,L is defined here as having a Vhalf negative to –55 mV. Kcna10+/+,+/– with gK,L, 17 ± 5 nS/pF (19); Kcna10+/+,+/– without gK,L, 3.7 ± 0.4 nS/pF (22); Kcna10–/–, 1.8 ± 0.4 nS/pF (13). Kcna10+/+,+/– with gK,L vs Kcna10–/–: p = 0.007, KWA, g 1.0. Asterisks: **p < 0.01. Line, median; Box, interquartile range; Whiskers, outliers.
Figure 2
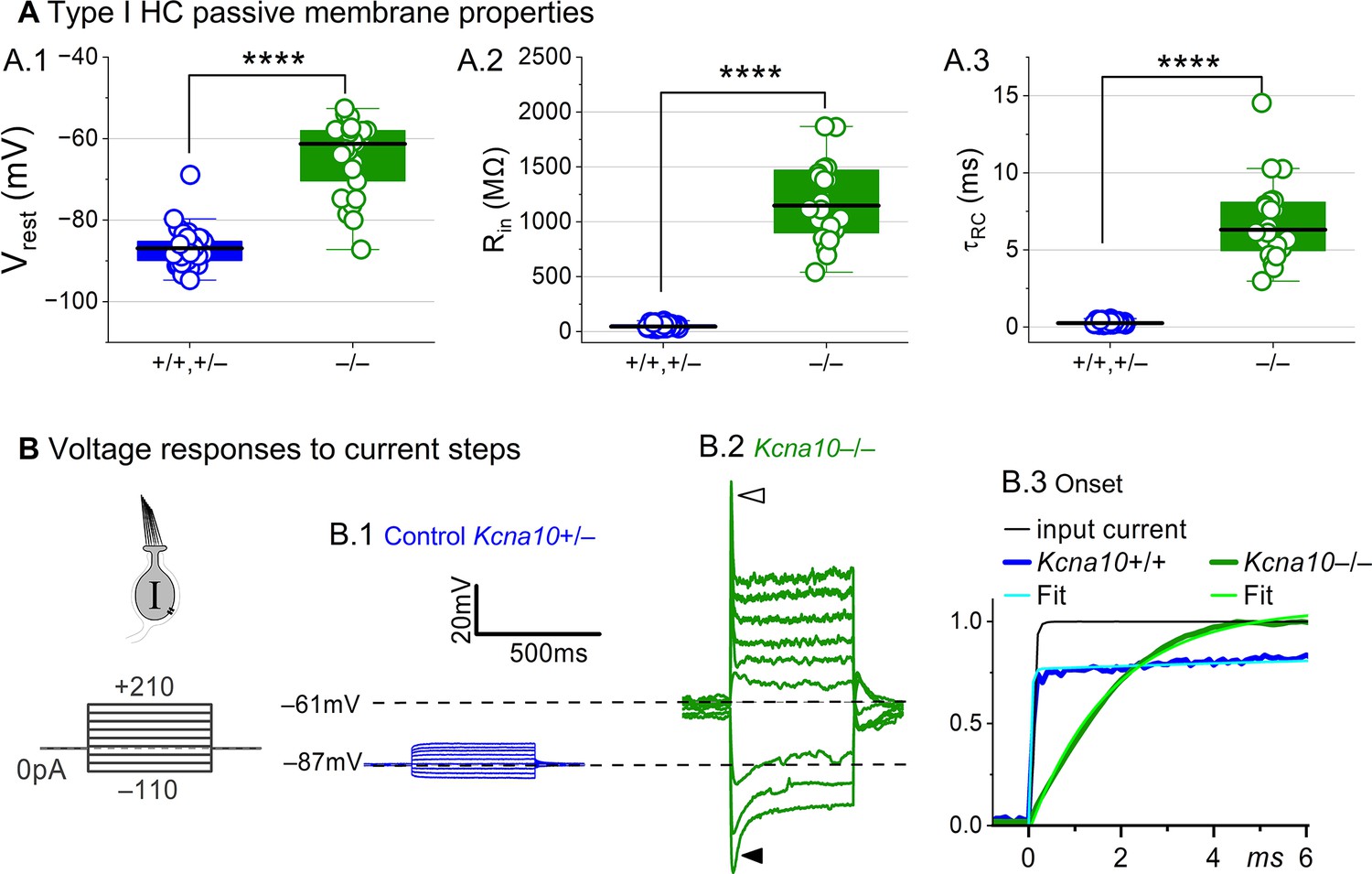
Kcna10–/– type I hair cells (HCs) had much longer membrane charging times and higher input resistances (voltage gains) than Kcna10+/+,+/– type I HCs.
(A) gK,L strongly affects passive membrane properties: (A.1) Vrest, Tukey’s test p<1E-9, (A.2) Rin, input resistance, Tukey’s test p<1E-9, and (A.3) membrane time constant, , Tukey’s test p<1E-9. See Table 2 for all statistics. (B) Current clamp responses to the same scale from (B.1) Kcna10+/– and (B.2) Kcna10–/– type I cells, both P29. Filled arrowhead (B.2), sag indicating IH activation. Open arrowhead, Depolarization rapidly decays as IDR activates. (B.3) First 6 ms of voltage responses to 170 pA injection, normalized to steady-state value; curves, double-exponential fits (Kcna10+/+, 40 μs and 2.4 ms) and single-exponential fits (Kcna10–/–, 1.1 ms). Asterisks, ****p < 0.0001. Line, median; Box, interquartile range; Whiskers, outliers.
Figure 3 with 3 supplements
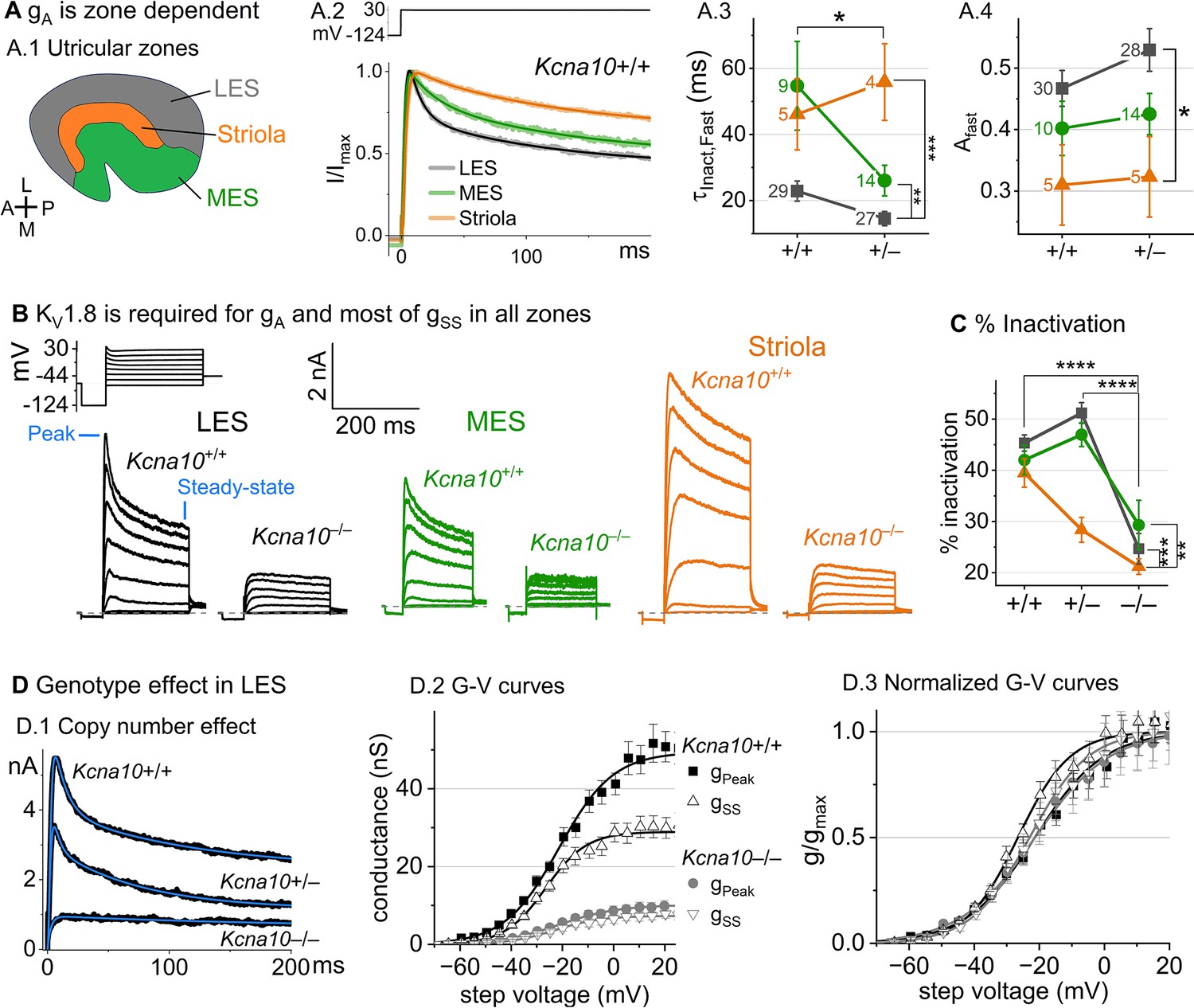
Kcna10–/– type II hair cells (HCs) in all zones of the sensory epithelium lacked the major rapidly inactivating conductance, gA, and had less delayed rectifier conductance.
Activation and inactivation varied with epithelial zone and genotype. (A) gA inactivation time course varied across zones. (A.1) Zones of the utricular epithelium: lateral extrastriola (LES), medial extrastriola (MES), and striola (S). (A.2) Normalized currents evoked by steps from –124 to +30 mV with overlaid fits of Equation 3. (A.3) was faster in Kcna10+/– (n=45) than Kcna10+/+ (n=43) HCs (KWA, p=0.027), and faster in LES (n=56) than MES (n=23, KWA, p=0.002) or S (n=9, KWA, p=2E-4). Point label is number of cells. Brackets show post hoc pairwise comparisons between two zones (vertical brackets) and horizontal brackets compare two genotypes; see Table 3 for statistics on kinetics. (A.4) Fast inactivation was a greater fraction of total inactivation in LES (n=58) than striola (n=10, Tukey’s test p=0.0041). (B) Exemplars; ages, left to right, P312, P53, P287, P49, P40, P154. (C) % inactivation at 30 mV was much lower in Kcna10–/– (n=37) than Kcna10+/– (n=47, Tukey’s HSD, p<1E-9) and Kcna10+/+ (n=44, Tukey’s HSD, p<1E-9). % inactivation was lower in striola (n=16) than LES (n=77, Tukey’s HSD, p=3E-5) and MES (n=36, Tukey’s HSD, p=0.0011). 2-way ANOVA detected interaction between zone and genotype, p=0.026 (Table 3). (D) Exemplar currents and G–V curves from LES type II HCs show a copy number effect. (D.1) Exemplar currents evoked by steps from –124 to +30 mV fit with Equation 3. (D.2) Averaged peak and steady-state conductance–voltage data points from LES cells (+/+, n=37; –/–, n=20) were fit with Boltzmann equations (Equation 1) and normalized by gmax in (D.3). Asterisks: *p < 0.05; **p < 0.01; ***p < 0.001; and ****p < 0.0001. Error bars, SEM. See Table 4 for statistics on voltage dependence.
Figure 3—figure supplement 1
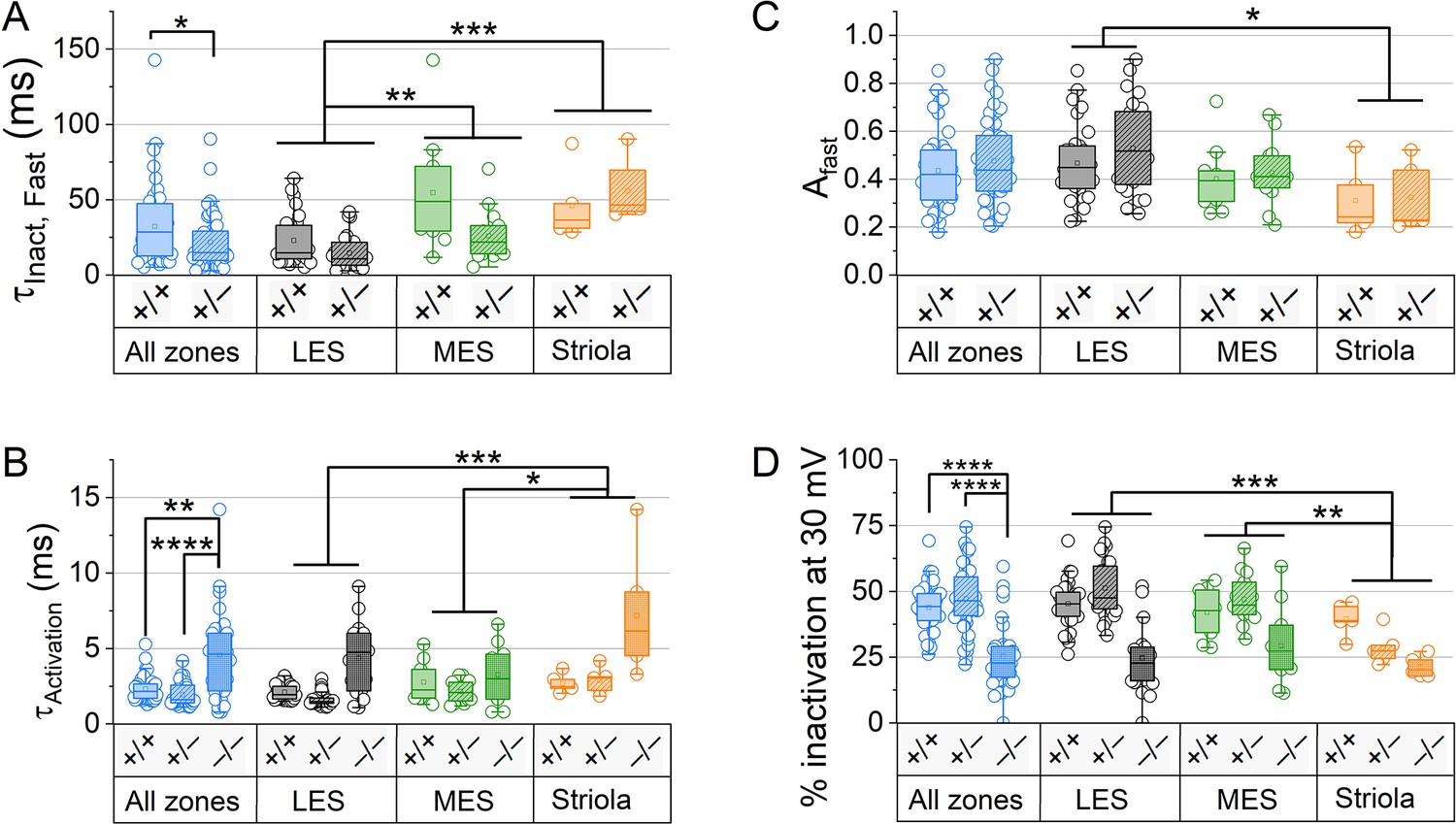
For type II hair cells (HCs) older than P12, KV conductance activation and inactivation differed across zones and genotypes.
(A) In Kcna10+/+ and Kcna10+/– HCs, at 30 mV was faster in LES (n=56) than MES (n=23, KWA p=0.002) or S (n=9, KWA p=2E-4). was faster in Kcna10+/– (n=45) than Kcna10+/+ (n=43, KWA p=0.027, see Table 3). (B) Fast inactivation was a larger fraction of total inactivation in LES (n=56) than striola (n=9, Tukey’s p=0.0041). (C) at 30 mV was slower in Kcna10–/– (n=53) than Kcna10+/+ (n=49, KWA p=0.0048) and Kcna10+/– (n=59, KWA p=2E-7), and slower in S (n=16) than LES (n=75, KWA p=6E-4) and MES (n=33, KWA p=0.02). (D) Percent inactivation at 30 mV was lower in S (n=20) than LES (n=99, 2-way ANOVA Tukey’s p<0.0001) and MES (n=42, 2-way ANOVA Tukey’s p=0.001), and lower in Kcna10–/– (n=43) than Kcna10+/+ (n=58, 2-way ANOVA Tukey’s p=0.0048 and <0.0001) and Kcna10+/– (n=60, 2-way ANOVA Tukey’s p<0.0001). Interaction between Zone and Genotype was significant (p=0.026). Asterisks: *p < 0.05; **p < 0.01; ***p < 0.001; and ****p < 0.0001. Line, median; Box, interquartile range; Whiskers, outliers.
Figure 3—figure supplement 2
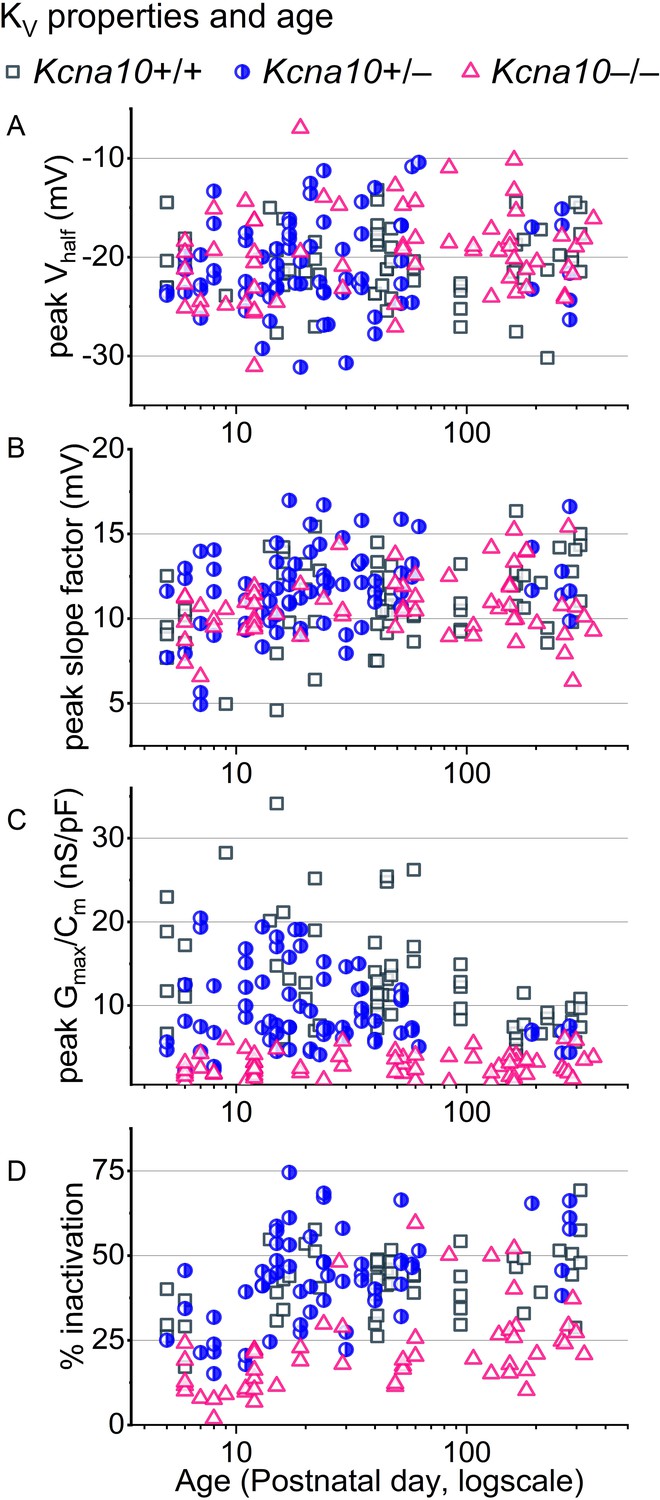
For type II hair cells (HCs) older than P12, KV conductances were stable.
(A–C) Parameters from Boltzmann fits of peak G–V relations and (D) % inactivation at +30 mV plotted against age from all zones. Overlaid curves are smoothing cubic β-splines. Note the seven extrastriolar Kcna10–/– type II HCs with % inactivation >30%.
Figure 3—figure supplement 3
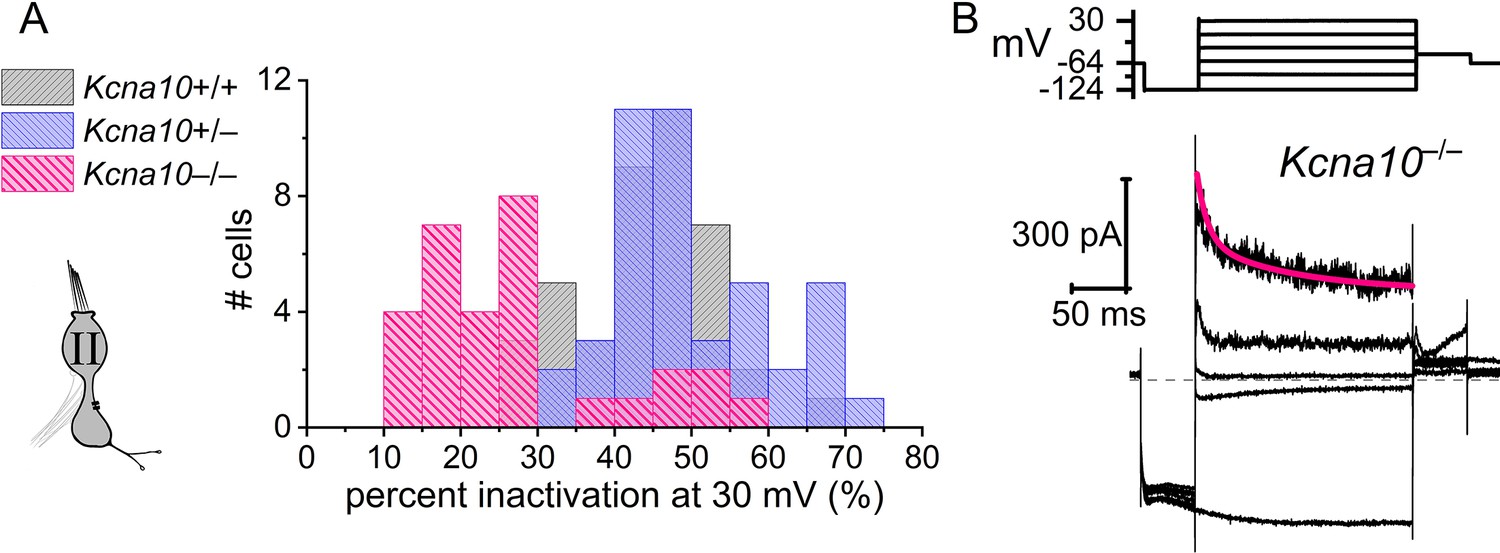
A minority of extrastriolar Kcna10–/– type II hair cells (HCs) had a very small fast-inactivating outward rectifier current.
(A) All extrastriolar Kcna10+/+,+/– type II HCs inactivated by >30%. Most mature (>P12) extrastriolar Kcna10–/– type II HCs inactivated by <30% but some inactivated by >30% (7/30, 23%) because they had fast inactivation (B). (B) Exemplar residual fast inactivation ( = 10 ms at +30 mV). For the seven cells in this group, = 30 ± 6 ms, amplitude of fast inactivation = 310 ± 70 pA; activation peak Vhalf = –15 ± 2 mV and slope factor = 12.4 ± 0.9 mV. These parameters are similar to gA but for the much smaller conductance (one-way ANOVAs).
Figure 4
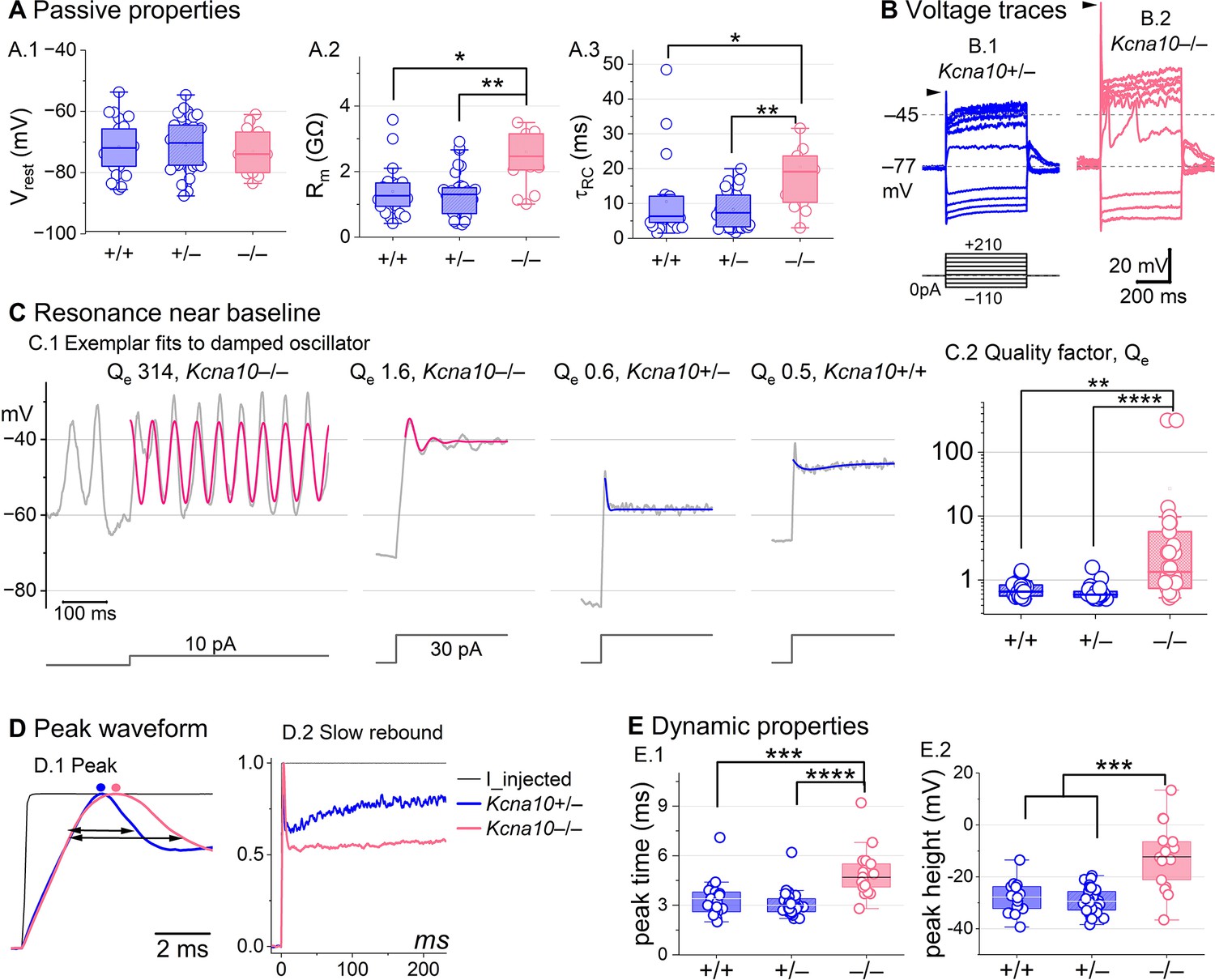
Kcna10–/– type II hair cells (HCs) had larger, slower voltage responses and more electrical resonance.
(A) Passive membrane properties near resting membrane potential: (A.1) Resting potential. Rinput (A.2) and (A.3) were obtained from single-exponential fits to voltage responses <15 mV. Rinput and were higher in Kcna10–/– (n=13) than Kcna10+/+ (n=22, KWA p=0.015; p=0.016) and Kcna10+/– (n=33, KWA p=0.002; p=0.008; see Table 5). (B) Exemplar voltage responses to iterated current steps (bottom) illustrate key changes in gain and resonance with KV1.8 knockout. (B.1) Kcna10+/– type II HC (P24, LES) and (B.2) Kcna10–/– type II HC (P53, LES). Arrowheads, depolarizing transients. (C) Range of resonance illustrated for Kcna10–/– type II HCs (left, pink curves fit to Equation 5) and controls (right, blue fits). (C.1) Resonant frequencies, left to right: 19.6, 18.4, 34.4, and 0.3 Hz. Leftmost cell resonated spontaneously (before step). (C.2) Tuning quality (Qe; Equation 6) was higher for Kcna10–/– (n=26) type II HCs (KWA: p = 0.0064 vs Kcna10+/+, n=23; p = 7E-8 vs Kcna10+/–, n=45). (D) Kcna10–/– type II HCs had higher, slower peaks and much slower rebound potentials in response to large (170 pA) current steps. (D.1) Normalized to show initial depolarizing transient (filled circles, times of peaks; horizontal arrows, peak width at half-maximum). (D.2) Longer time scale to highlight how null mutation reduced post-transient rebound. (E) In Kcna10–/– HCs (n=19), depolarizing transients evoked by a +90 pA step were slower to peak (E.1) than in Kcna10+/+ (n=19, 2-way ANOVA Tukey’s p<1E-9) and Kcna10+/– (n=34, 2-way ANOVA Tukey’s p<1E-9) and (E.2) larger than in Kcna10+/+ (n=19, KWA p=0.006) and Kcna10+/– (n=34, KWA p=2E-4). Asterisks: *p < 0.05; **p < 0.01; ***p < 0.001; and ****p < 0.0001. Line, median; Box, interquartile range; Whiskers, outliers.
Figure 5
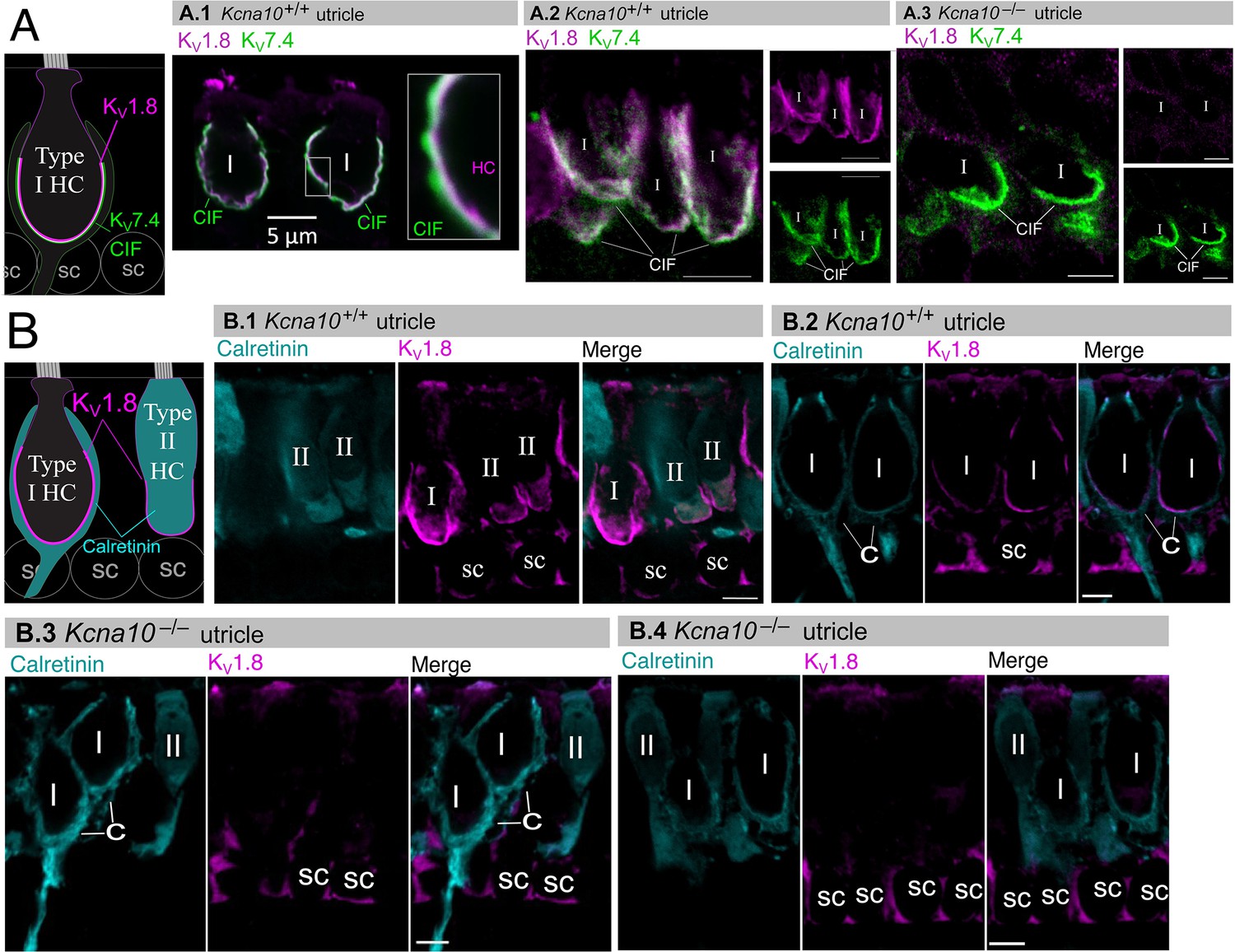
Type I and II hair cell (HC) basolateral membranes show specific immunoreactivity to Kv1.8 antibody (magenta).
Antibodies for KV7.4 (A, green) and calretinin (B, cyan) were used as counterstains for calyx membrane (Kv7.4), type II HC cytoplasm (calretinin) and cytoplasm of striolar calyx-only afferents (calretinin). (A) Left, Cartoon showing KV7.4 on the calyx inner face membrane (CIF) and KV1.8 on the type I HC membrane. SC, supporting cell nuclei. A.1–3, Adult mouse utricle sections. KV7.4 antibody labeled calyces on two KV1.8-positive type I HCs (A.1), four KV1.8-positive type I HCs (A.2), and two KV1.8-negative type I HCs from a Kcna10–/– mouse (A.3). (B) Left, Cartoon showing cytoplasmic calretinin stain in calyx-only striolar afferents and most type II HCs, and KV1.8 on membranes of both HC types. In wildtype utricles, KV1.8 immunolocalized to basolateral membranes of type I and II HCs (extrastriola, B.1). KV1.8 immunolocalized to type I HCs (striola, B.2). Staining of supporting cell (SC) membranes by Kv1.8 antibody was non-specific, as it was present in Kcna10–/– tissue (striola, B.3 and B.4). All scale bars 5 µm.
Figure 6
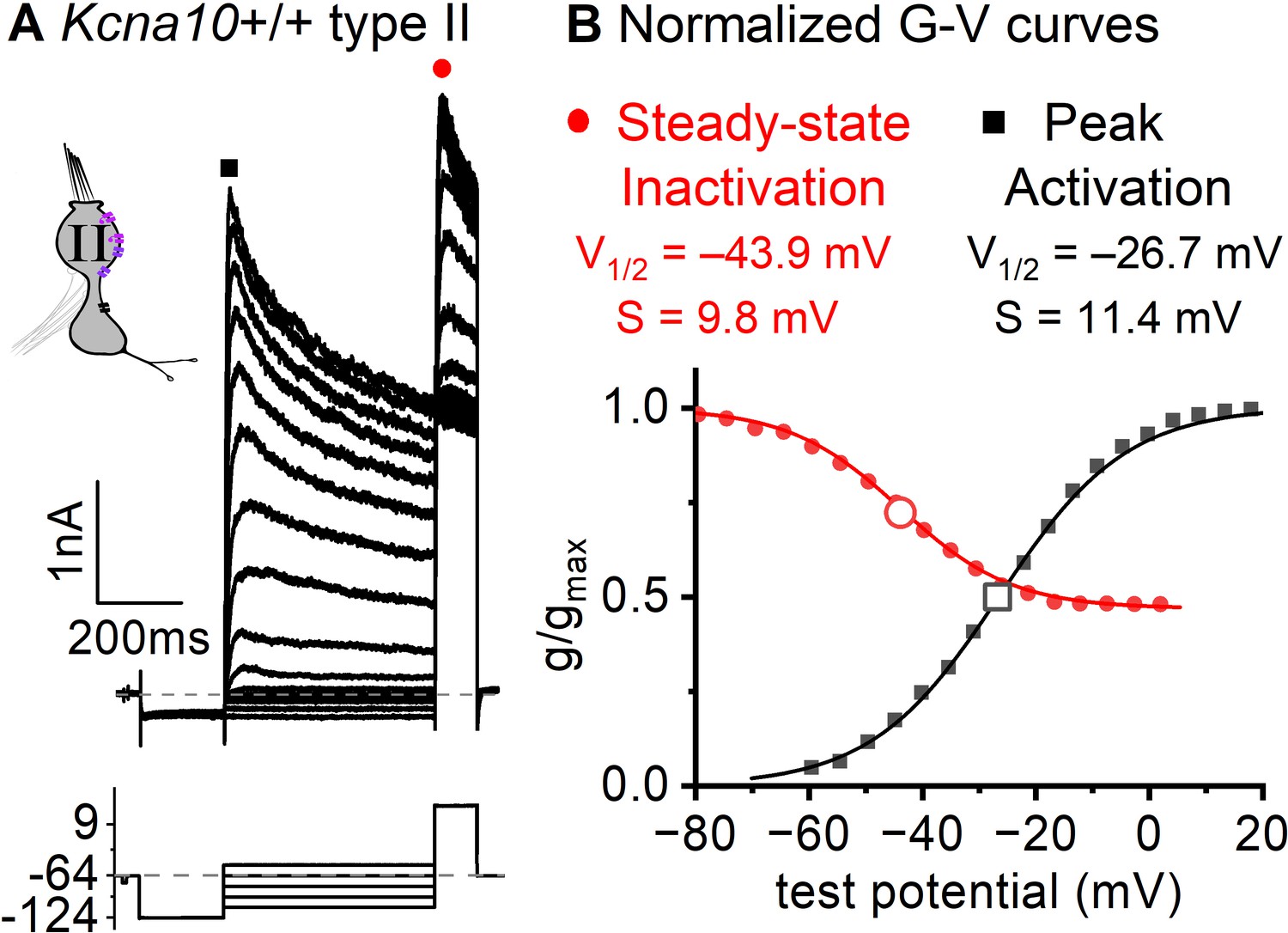
Inactivation curve of gA in extrastriolar type II hair cells (HCs).
(A) Modified voltage protocol measured accumulated steady-state inactivation at the tail potential. 100 μM ZD7288 in bath prevented contamination by HCN current. (B) Voltage dependence of gA’s steady-state inactivation (h∞ curve) and peak activation are consistent with KV1.4 heteromers. Curves, Boltzmann fits (Equation 1). Average fit parameters from Kcna10+/+,+/– type II HCs, P40–P210, median P94. Inactivation: Vhalf, –42 ± 2 mV (n = 11); S, 11 ± 1 mV. Activation: Vhalf, –23 ± 1 mV (n = 11); S, 11.2 ± 0.4 mV.
Figure 7 with 2 supplements
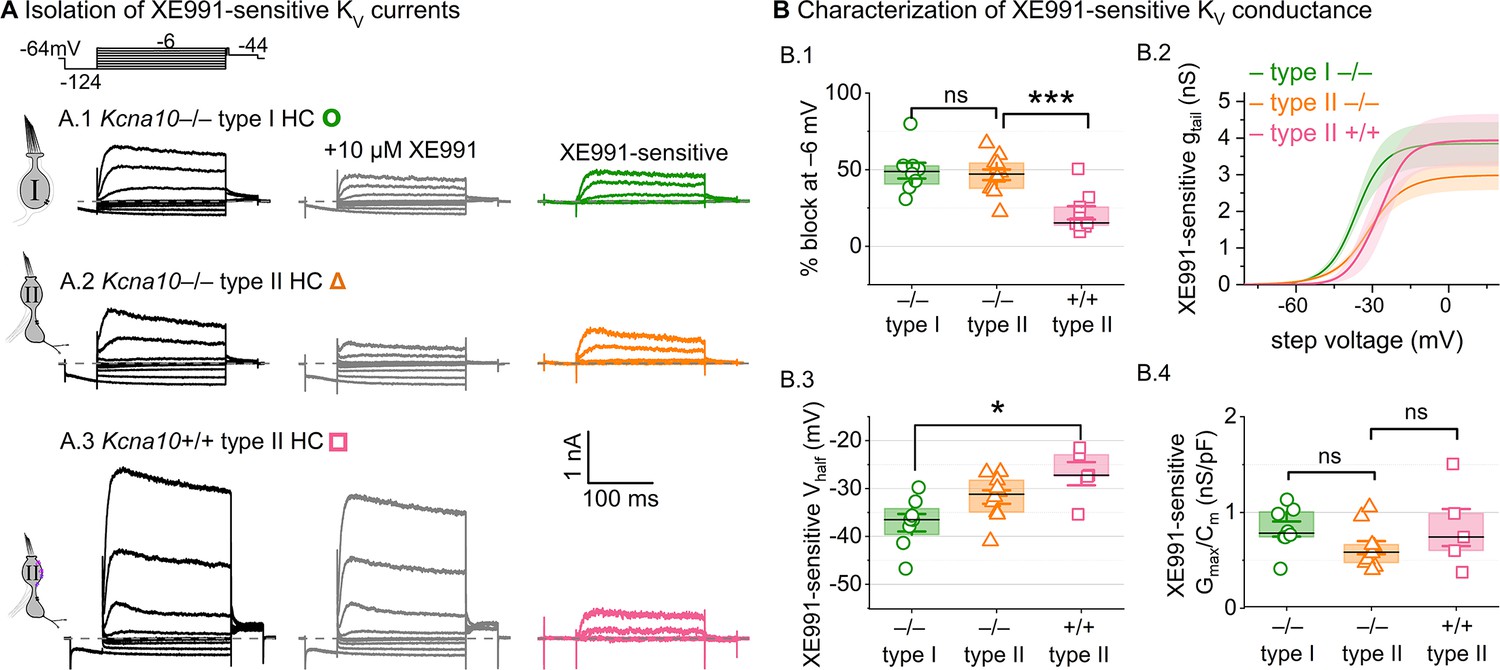
A KV7-selective blocker, XE991, reduced residual delayed rectifier currents in Kcna10–/– type I and II hair cells (HCs).
(A) XE991 (10 μM) partly blocked similar delayed rectifier currents in type I and II Kcna10–/– HCs and a type II Kcna10+/+ HC. (B) Properties of XE991-sensitive conductance, DR(KV7). (B.1) % Block of steady-state current. (B.2) Mean tail G–V curves for Kcna10–/– type I HCs (n = 8), Kcna10–/– type II HCs (9), and Kcna10+/+ type II HCs (5); shading is ± SEM. (B.3) Vhalf was less negative in Kcna10+/+ type II than Kcna10–/– type I HC (p = 0.01, KWA). (B.4) Conductance density was similar in all groups (ANOVA), non-significant at 0.4 power (left), 0.2 power (right). Asterisks: *p < 0.05 and ***p < 0.001. Line, median; Box, interquartile range; Whiskers, outliers.
Figure 7—figure supplement 1
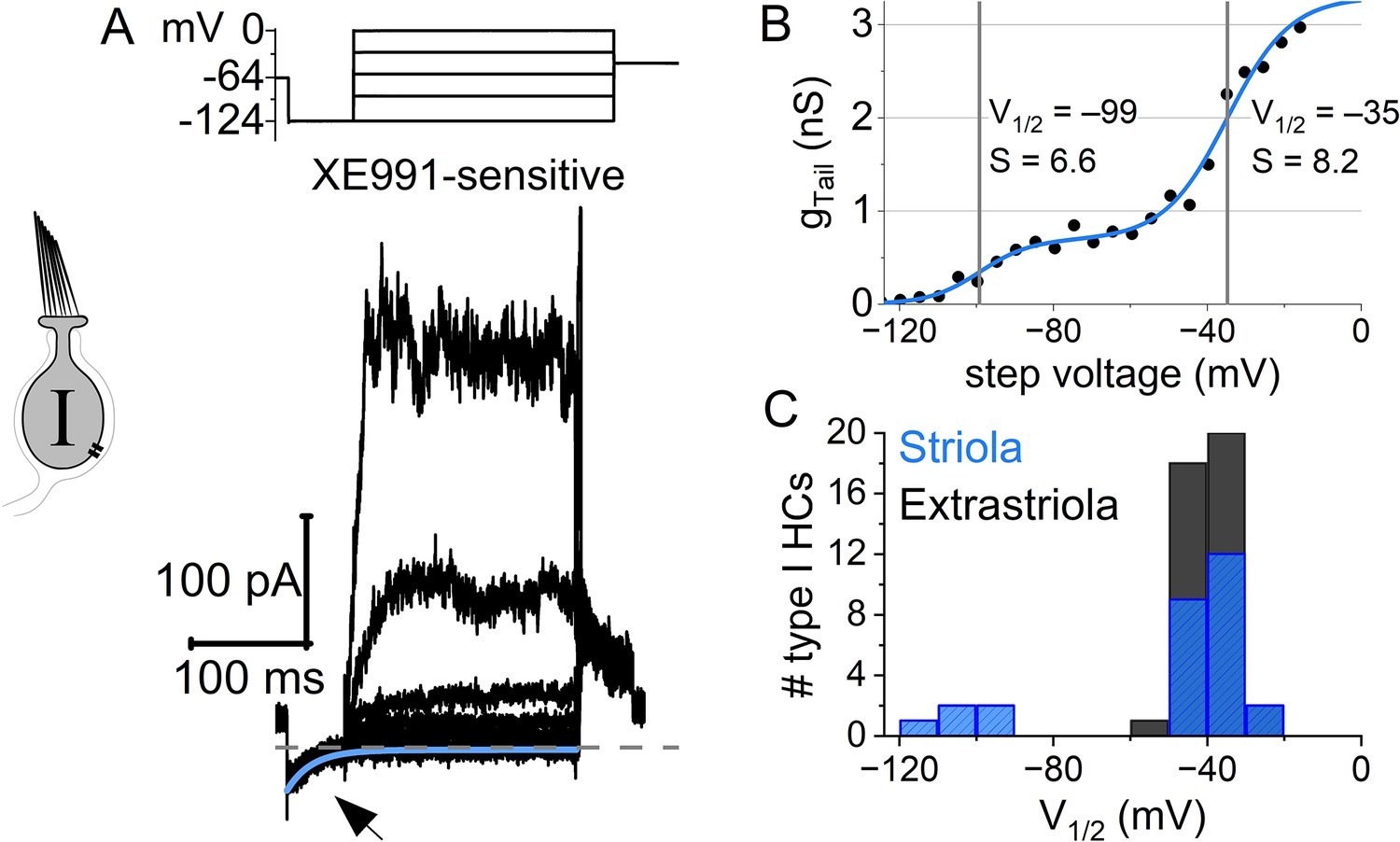
A minority of striolar Kcna10–/– type I hair cells (HCs) had a small low-voltage-activated outward rectifier current in addition to a more positively activating outward rectifier.
(A) Low-voltage-activated current from one cell was isolated by subtraction with 10 μM XE991 (P39), indicating that it was carried by KV7 channels. Deactivation of XE991-sensitive current evoked by step from –64 to –124 mV (arrow) was fit with exponential decay ( = 21 ms). (B) XE991-sensitive tail G–V curve of the XE991-blocked conductance (A) was fit with a sum of two Boltzmann equations: G(V) = A1/(1 + exp((Vhalf,1 – V)/S1)) + A2/(1 + exp((Vhalf,2 – V)/S2)). (C) The low-voltage-activated Vhalf,1 component was only seen in striolar Kcna10–/– type I HCs, and even there in the minority: 5/23; 22%; P6–P370. It was always seen together with a more positively activating outward rectifier. Average Boltzmann parameters (n=5, including B): A1/(A1 + A2) = 0.15 ± 0.04, Vhalf,1 = –106 ± 5 mV, S1 = 3.8 ± 0.8 mV, Vhalf,2 = –41±1 mV, S2 = 7 ± 1 mV. Ages: P11, 39, 202, 202, 202. No extrastriolar type I HCs (0/45; P6–277) had a double activation tail G–V curve.
Figure 7—figure supplement 2
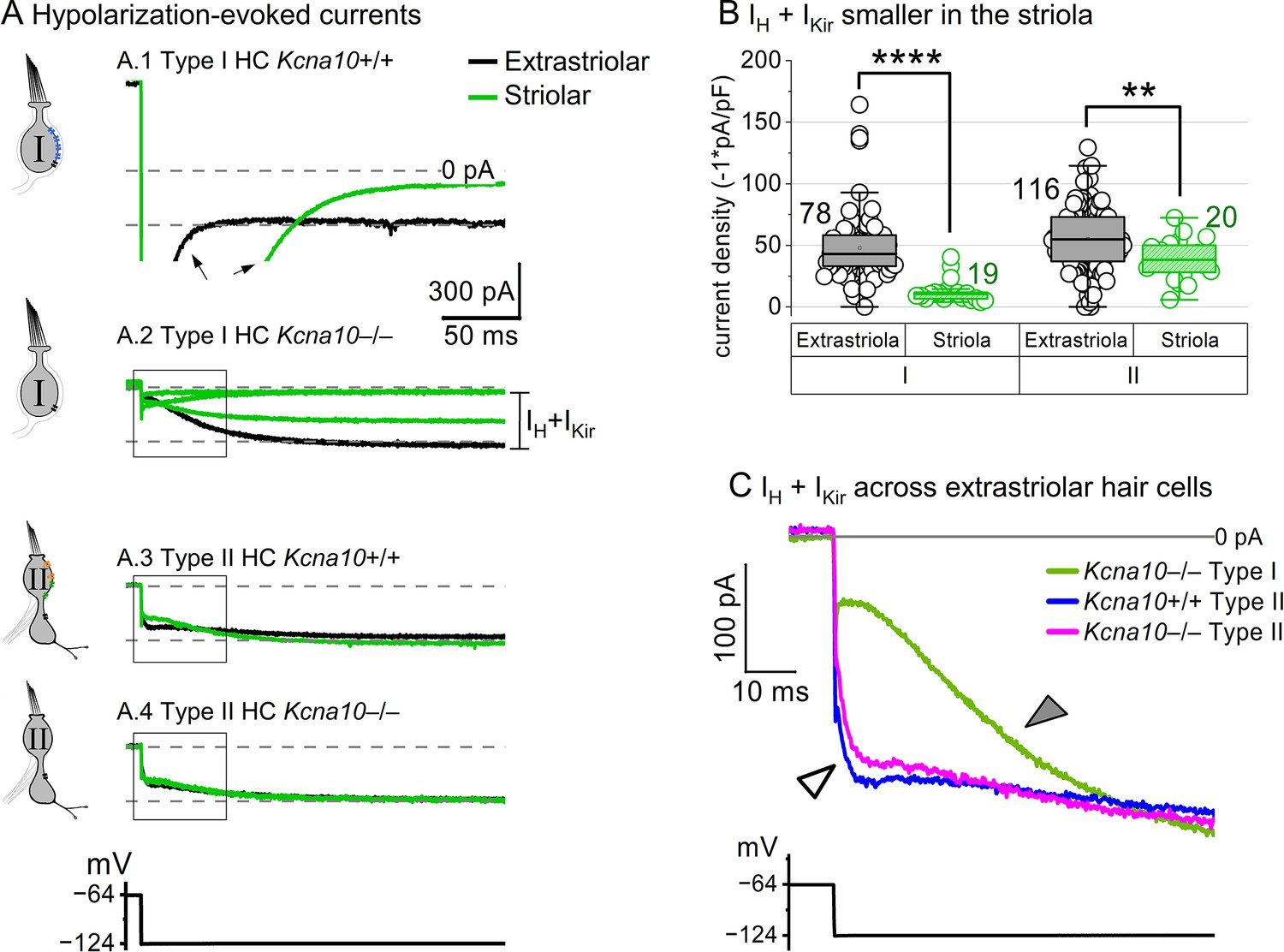
No difference was detected in H (HCN) and Kir (fast inward rectifier) currents between Kcna10+/+ and Kcna10–/– hair cells (HCs), consistent with a specific involvement of KV1.8 in Kcna10 expression.
(A) Hyperpolarizing voltage steps evoked IKir and IHCN in Kcna10+/+,+/–,–/– type I and II HCs. (A.1) Arrows, deactivation of gK,L. (A.2–A.4) Bracket, the sum of IH and IKir were measured as total inward current after 250 ms at –124 mV. (B) Summed IKIR and IH density was smaller in striola than extrastriola (type I HC KWA, p=4E-9; type II HC 2-way ANOVA Tukey’s p=0.006, see Supplementary file 1c). Data labels are number of cells. (C) Inward currents seen at the onset of hyperpolarization, including IKir, were larger in type II HCs than type I. Magnification of boxed in inset from the extrastriolar cells in A.2–A.4. Open arrowhead, activation of fast inward rectifier, IKir; filled arrowhead, slower activation of IHCN. Asterisks: **p < 0.01 and ****p < 0.0001. Line, median; Box, interquartile range; Whiskers, outliers.
Tables
Table 1
Type I hair cell KV activation voltage dependence.
Mean ± SEM (number of cells). g is effect size, Hedge’s g. KWA is Kruskal–Wallis ANOVA.
| Zone | Kcna10 | Tail V1/2, mV* | Tail S, mV† | Tail gmax, nS‡ | Tail gmax/Cm, nS/pF§ | Age (median, range) |
|---|---|---|---|---|---|---|
| Extrastriola | +/+ | –85 ± 2 (12) | 4.3 ± 0.4 (12) | 270 ± 40 (11) | 47 ± 8 (11) | 22, 14–287 |
| +/– | –83 ± 1 (40) | 5.2 ± 0.3 (40) | 210 ± 20 (40) | 37 ± 4 (40) | 19, 13–259 | |
| –/– | –40.2 ± 0.9 (26) | 5.7 ± 0.3 (26) | 5.4 ± 0.3 (26) | 1.11 ± 0.08 (26) | 45, 14–277 | |
| Striola | +/+ | –87 ± 3 (6) | 4.3 ± 0.3 (6) | 310 ± 70 (6) | 41 ± 7 (6) | 40, 15–59 |
| +/– | –88 ± 2 (3) | 4.7 ± 0.9 (3) | 270 ± 60 (3) | 44 ± 6 (3) | 19, 14–20 | |
| –/– | –38 ± 1 (13) | 6.2 ± 0.4 (13) | 6.5 ± 0.6 (13) | 1.5 ± 0.1 (13) | 202, 14–370 |
-
*
–/– vs +/+: two-way ANOVA, p < 1E−9, g 7.7; –/– vs +/–: two-way ANOVA, p < 1E−9, g 6.8.
-
†
–/– vs +/+: two-way ANOVA, p = 8.4E−4, g 1.2.
-
‡
–/– vs +/+: two-way ANOVA, p < 1E−9, g 3.7; –/– vs +/–: two-way ANOVA, p < 1E−9, g 2.1.
-
§
–/– vs +/+: two-way ANOVA, p < 1E−9, g 3.6; –/– vs +/–: two-way ANOVA, p < 1E−9, g 2.0.
Table 2
Type I hair cell passive membrane properties in the extrastriola (ES) and striola (S).
Mean ± SEM (number of cells). g is effect size, Hedge’s g. KWA is Kruskal–Wallis ANOVA.
| Zone | Kcna10 | Vrest, mV*, † | Rinput, MΩ‡ | , ms§ | Cm, pF¶ | Age (median, range) |
|---|---|---|---|---|---|---|
| ES | +/+ | –84 ± 3 (6) | 44 ± 6 (6) | 0.24 ± 0.03 (6) | 6.1 ± 0.4 (13) | 20, 14–287 |
| +/– | –88.0 ± 0.7 (28) | 55 ± 5 (24) | 0.32 ± 0.03 (23) | 5.8 ± 0.2 (44) | 21, 16–29 | |
| –/– | –63 ± 2 (15) | 1400 ± 100 (15) | 6.4 ± 0.6 (15) | 5.0 ± 0.2 (27) | 45, 14–202 | |
| S | +/+ | –87 ± 2 (4) | 50 ± 8 (4) | 0.30 ± 0.04 (4) | 7.4 ± 0.7 (7) | 43, 40–59 |
| +/– | –87 ± 3 (3) | 38 ± 8 (2) | 0.21 ± 0.01 (2) | 5.9 ± 0.6 (3) | 19, 19–20 | |
| –/– | –74 ± 5 (5) | 1000 ± 300 (4) | 4.2 ± 1.0 (4) | 4.4 ± 0.2 (14) | 202, 24–370 |
-
*
Striolar –/– vs ES –/–: two-way ANOVA, p = 0.006, g 1.2; striolar –/– vs striolar +/+,+/–: two-way ANOVA, p = 0.005, g 1.7.
-
†
–/– vs +/+: two-way ANOVA, p < 1E−9, g 2.3; –/– vs +/–: two-way ANOVA, p < 1E−9, g 3.4.
-
‡
–/– vs +/+: two-way ANOVA, p < 1E−9, g 3.1; –/– vs +/–: two-way ANOVA, p < 1E−9, g 3.9.
-
§†
–/– vs +/+: two-way ANOVA, p < 1E−9, g 2.7; –/– vs +/–: two-way ANOVA, p < 1E−9, g 3.4.
-
¶‡
–/– vs +/+: two-way ANOVA, p = 3E−7, g 1.5; –/– vs +/–: two-way ANOVA, p = 1.3E−4, g 1.0; +/–vs +/+: two-way ANOVA, p = 0.048, g 0.6.
Table 3
Type II hair cell KV currents: activation and inactivation time course at +30 mV.
Mean ± SEM. g is effect size, Hedge’s g. KWA is Kruskal–Wallis ANOVA.
| Zone | Kcna10 | at 30 mV, ms*, † | at 30 mV, ms ‡, § | Fast inactivation prominence¶ | Inactivation %**,†† | N cells | Age (median, range) |
|---|---|---|---|---|---|---|---|
| LES | +/+ | 2.11 ± 0.09 | 23 ± 3 | 0.46 ± 0.03 | 45 ± 2 | 30 | 46, 14–312 |
| +/– | 1.64 ± 0.09 | 15 ± 2 | 0.53 ± 0.03 | 51 ± 2 | 27 | 29, 13–280 | |
| –/– | 4.4 ± 0.5 | NA | NA | 25 ± 3 | 21 | 128, 15–355 | |
| MES | +/+ | 2.8 ± 0.5 | 50 ± 10 | 0.40 ± 0.04 | 42 ± 3 | 9 | 94, 22–296 |
| +/– | 2.2 ± 0.2 | 90 ± 60 | 0.42 ± 0.03 | 47 ± 2 | 15 | 24, 13–52 | |
| –/– | 10 ± 7 | NA | NA | 29 ± 5 | 10 | 84, 28–355 | |
| Striola | +/+ | 2.7 ± 0.3 | 50 ± 10 | 0.31 ± 0.07 | 39 ± 3 | 5 | 45, 40–287 |
| +/– | 2.9 ± 0.4 | 300 ± 200 | 0.3 ± 0.06 | 28 ± 2 | 5 | 19, 14–30 | |
| –/– | 7 ± 2 | NA | NA | 22 ± 2 | 6 | 202, 29–298 |
-
*
–/– vs +/+: KWA, p = 0.0048, g 0.6; –/– vs +/–: KWA, p = 2.3E−7, g 0.6.
-
†
Striola vs LES: KWA, p = 5.7E−4, g 1.0.
-
‡
+/– vs +/+: KWA, p = 0.027, g 0.2.
-
§
LES vs MES: KWA, p = 0.0018, g 0.3; LES vs Striola: KWA, p = 1.9E−4, g 0.8.
-
¶
LES vs Striola: two-way ANOVA, p = 0.0041, g 0.7.
-
**
–/– vs +/+: two-way ANOVA, p < 1E−9, g 1.7; –/– vs +/–: two-way ANOVA, p < 1E−9, g 1.8.
-
††
Striola vs LES: two-way ANOVA, p = 3.4E−5, g 0.9; Striola vs MES: two-way ANOVA, p = 0.0011, g 1.0; interaction between genotype and zone: two-way ANOVA, p = 0.026.
Table 4
Type II hair cell KV currents: activation voltage dependence.
Mean ± SEM. g is effect size, Hedge’s g. KWA is Kruskal–Wallis ANOVA.
| Zone | Kcna10 | Peak V1/2, mV** | Peak S, mV††, ‡ | A-type gmax/Cm, nS/pF§ § | SS Vhalf, mV¶ ¶ | SS S, mV**** | SS gmax/Cm, nS/pF †† ‡ ‡ | N cells | Age (median, range) |
|---|---|---|---|---|---|---|---|---|---|
| LES | +/+ | –19.8 ± 0.6 | 11.8 ± 0.4 | 4.0 ± 0.3 | –25.0 ± 0.5 | 8.7 ± 0.3 | 7.1 ± 0.8 | 37 | 46, 14–312 |
| +/– | –19.8 ± 0.8 | 12.8 ± 0.4 | 3.8 ± 0.3 | –26.8 ± 0.8 | 8.7 ± 0.3 | 4.9 ±0.4 | 35 | 29, 13–280 | |
| –/– | –18 ± 1 | 11.7 ± 0.4 | 0.37 ± 0.05 | –19 ± 1 | 12.1 ± 0.5 | 1.8 ±0.2 | 20 | 128, 15–355 | |
| MES | +/+ | –22 ± 1 | 11 ± 0.7 | 4.1 ± 0.7 | –26 ± 1 | 8.3 ± 0.5 | 9 ±1 | 11 | 94, 22–296 |
| +/– | –21 ± 1 | 11.8 ± 0.4 | 3.6 ± 0.5 | –27 ± 1 | 9.0 ± 0.3 | 5.9 ±0.7 | 16 | 24, 13–52 | |
| –/– | –19 ± 1 | 10.8 ± 0.6 | 0.6 ± 0.1 | –20 ± 1 | 10.7 ± 0.7 | 2.5 ±0.3 | 15 | 84, 28–355 | |
| Striola | +/+ | –24 ± 1 | 9.6 ± 0.5 | 5 ± 1 | –26.6 ± 0.9 | 8.2 ± 0.4 | 12 ±1 | 7 | 45, 40–287 |
| +/– | –25 ± 2 | 9.4 ± 0.4 | 2.6 ± 0.6 | –28 ± 2 | 8.2 ± 0.3 | 10±2 | 6 | 19, 14–30 | |
| –/– | –21.3 ± 0.9 | 10.3 ± 0.5 | 0.7 ± 0.1 | –21.7 ± 0.8 | 10.5 ± 0.6 | 3.9±0.5 | 8 | 202, 29–298 |
-
*
Striola vs LES: two-way ANOVA, p = 0.00116, g 0.9.
-
†
Striola vs MES: two-way ANOVA, p = 0.016, g 0.8; Striola vs LES: two-way ANOVA, p = 7.5E−6, g 1.2.
-
‡
–/– vs +/–: two-way ANOVA, p = 0.036, g 0.5.
-
§
–/– vs +/+: Welch ANOVA, p < 1E−9, g 2.3; –/– vs +/–: Welch ANOVA, p < 1E−9, g 2.3.
-
¶
–/– vs +/+: two-way ANOVA, p < 1E−9, g 1.4; –/– vs +/–: two-way ANOVA, p < 1E−9, g 1.6.
-
**
–/– vs +/+: two-way ANOVA, p < 1E−9, g 1.4; –/– vs +/–: two-way ANOVA, p = 4.5E−7, g 1.1.
-
††
–/– vs +/+: Welch ANOVA, p < 1E−9, g 1.6; –/– vs +/–: Welch ANOVA, p < 1E−9, g 1.3; +/+vs +/–: Welch ANOVA, p = 0.007, g 1.6.
-
‡ ‡
Striola vs LES: one-way ANOVA, p = 0.001, g (0.9); Striola vs MES: one-way ANOVA, p = 0.01, g 0.8.
Table 5
Type II hair cell passive membrane properties in the extrastriola (ES) and striola (S).
Mean ± SEM (number of cells). g is effect size, Hedge’s g. KWA is Kruskal–Wallis ANOVA. Peak height and time were measured from responses to 170 pA input from rest.
| Zone | Kcna10 | Vrest, mV | Rinput, GΩ* | , ms† | Peak height, mV‡ | Peak time, ms§ | Cm, pF | Age (median, range) |
|---|---|---|---|---|---|---|---|---|
| ES | +/+ | –71 ± 2 (19) | 1.4 ± 0.2 (16) | 11 ± 3 (16) | –20 ± 2 (15) | 2.5 ± 0.2 (15) | 4.7 ± 0.2 (50) | 45, 16–312 |
| +/– | –71 ± 2 (34) | 1.2 ± 0.1 (27) | 9 ± 1 (27) | –20 ± 1 (30) | 2.44 ± 0.08 (30) | 4.6 ± 0.1 (52) | 27, 13–280 | |
| –/– | –76 ± 2 (9) | 2.3 ± 0.3 (7) | 16 ± 3 (7) | 2 ± 6 (7) | 3.6 ± 0.3 (7) | 4.6 ± 0.2 (35) | 53, 15–154 | |
| S | +/+ | –73.1 ± 1.0 (6) | 1.4 ± 0.1 (6) | 9 ± 1 (6) | –20 ± 2 (5) | 2.7 ± 0.1 (5) | 4.6 ± 0.2 (7) | 45, 40–224 |
| +/– | –71 ± 3 (5) | 1.4 ± 0.3 (6) | 7 ± 2 (6) | –20 ± 2 (6) | 2.3 ± 0.1 (6) | 4.8 ± 0.2 (6) | 19, 19–30 | |
| –/– | –68 ± 2 (6) | 3.0 ± 0.7 (6) | 26 ± 10 (6) | 2 ± 7 (4) | 4 ± 1 (4) | 4.4 ± 0.3 (7) | 178, 29–298 |
-
*
–/– vs +/+: KWA, p = 0.015, g 1.2; –/– vs +/–: KWA, p = 0.002, g 1.5.
-
†
–/– vs +/+: KWA, p = 0.016, g 0.7; –/– vs +/–: KWA, p = 0.008, g 1.2.
-
‡
–/– vs +/+: KWA, p = 0.006, g 2.1; –/– vs +/–: KWA, p = 2E−4, g 2.6.
-
§
–/– vs +/+: two-way ANOVA, p < 1E−9, g 1.3; –/– vs +/–: two-way ANOVA, p < 1E−9, g 1.9.
Key resources table
| Reagent type (species) or resource | Designation | Source or reference | Identifiers | Additional information |
|---|---|---|---|---|
| Antibody | Anti-Kv1.8 (Rabbit polyclonal) | Alomone | Cat# APC-157, lot# 0102, RRID:AB_2341039 | 1:200 or 1:400 |
| Antibody | Anti-calretinin (goat polyclonal) | Millipore | Cat# AB1550, lot# 9669, RRID:AB_90764 | 1:600 |
| Antibody | Anti-Kv7.4 (mouse IgG1 monoclonal) | NeuroMab | Cat# 2HK-65, RRID:AB_2131828 | 1:200 |
| Peptide, recombinant protein | Iberiotoxin | Alomone | STI-400 | 100 nM (water) |
| Chemical compound, drug | XE991 | Sigma | X2254 | 100 µM (water) |
| Chemical compound, drug | ZD7288 | Tocris | APN18035-2 | 100 µM (water) |
| Peptide, recombinant protein | Bovine serum albumin | Fisher | BP671 | 1 mg/ml (water) |
Additional files
-
Supplementary file 1
This file contains descriptive and comparative statistics on three additional analyses of the dataset.
(a) Test of sex differences in hair cell KV channel data. (b) We did not detect a genotype effect on soma size of type I hair cells (HCs). (c) IKir and IH were greater in the extrastriola (ES) than striola (S), but did not vary by genotype.
- https://cdn.elifesciences.org/articles/94342/elife-94342-supp1-v1.docx
-
MDAR checklist
- https://cdn.elifesciences.org/articles/94342/elife-94342-mdarchecklist1-v1.pdf
-
Source code 1
Fitkin.m is a MATLAB function to fit the activation and inactivation kinetics of KV currents to Equations 2; 3.
Fitkin.m processed current–time traces and output parameters found in Figure 3, Figure 3—figure supplement 1, and Table 3.
- https://cdn.elifesciences.org/articles/94342/elife-94342-code1-v1.zip
-
Source code 2
Fitzmann.m is a MATLAB function to fit the activation voltage dependence of KV currents to Equation 1.
Fitzmann.m processed G–V data points and output parameters found in Figure 1, Figure 3, Figure 7, Figure 1—figure supplement 1, Figure 3—figure supplement 2, Table 1, Table 4, and Supplementary file 1a.
- https://cdn.elifesciences.org/articles/94342/elife-94342-code2-v1.zip
Download links
A two-part list of links to download the article, or parts of the article, in various formats.
Downloads (link to download the article as PDF)
Open citations (links to open the citations from this article in various online reference manager services)
Cite this article (links to download the citations from this article in formats compatible with various reference manager tools)
The potassium channel subunit KV1.8 (Kcna10) is essential for the distinctive outwardly rectifying conductances of type I and II vestibular hair cells
eLife 13:RP94342.
https://doi.org/10.7554/eLife.94342.4




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}