Long-term hematopoietic transfer of the anti-cancer and lifespan-extending capabilities of a genetically engineered blood system by transplantation of bone marrow mononuclear cells
- The Ph.D. Program in Medicine Neuroscience, Taipei Medical University, Taiwan
- Institute of Molecular Biology, Academia Sinica, Taiwan
- Chang Gung Memorial Hospital, Taiwan
- Pro-Clintech Co. Ltd, Taiwan
- Department of Nursing, Chang Gung University of Science and Technology, Taiwan
- Community Medicine Research Center, Chang Gung Memorial Hospital, Keelung Branch, Taiwan
- Agricultural Biotechnology Research Center, Academia Sinica, Taiwan
- Graduate Institute of Life Sciences, National Defense Medical Center, Taiwan
- Genomics Research Center, Academia Sinica, Taiwan
eLife assessment
This useful article focuses on understanding how an Eklf mutation confers anti-cancer and longevity advantages in vivo. The data demonstrate that Eklf (K74R) imparts such advantages in a background- and age-independent manner in both female and male mice, and that the benefits are transferable by bone marrow transplantation. Despite added data since a previous version, the article unfortunately remains incomplete as it is still unclear whether Eklf affects resistance to malignant progression/metastasis by modulating Pd1 or Pdl1 or by increasing NK cells. The authors provide evidence that supports in principle both mechanisms, and they do not resolve which mechanism is primarily involved.
https://doi.org/10.7554/eLife.88275.3.sa0Significance of the findings:
Useful: Findings that have focused importance and scope
- Landmark
- Fundamental
- Important
- Valuable
- Useful
Strength of evidence:
Incomplete: Main claims are only partially supported
- Exceptional
- Compelling
- Convincing
- Solid
- Incomplete
- Inadequate
During the peer-review process the editor and reviewers write an eLife Assessment that summarises the significance of the findings reported in the article (on a scale ranging from landmark to useful) and the strength of the evidence (on a scale ranging from exceptional to inadequate). Learn more about eLife Assessments
Abstract
A causal relationship exists among the aging process, organ decay and disfunction, and the occurrence of various diseases including cancer. A genetically engineered mouse model, termed Klf1K74R/K74R or Klf1(K74R), carrying mutation on the well-conserved sumoylation site of the hematopoietic transcription factor KLF1/EKLF has been generated that possesses extended lifespan and healthy characteristics, including cancer resistance. We show that the healthy longevity characteristics of the Klf1(K74R) mice, as exemplified by their higher anti-cancer capability, are likely gender-, age-, and genetic background-independent. Significantly, the anti-cancer capability, in particular that against melanoma as well as hepatocellular carcinoma, and lifespan-extending property of Klf1(K74R) mice, could be transferred to wild-type mice via transplantation of their bone marrow mononuclear cells at a young age of the latter. Furthermore, NK(K74R) cells carry higher in vitro cancer cell-killing ability than wild-type NK cells. Targeted/global gene expression profiling analysis has identified changes in the expression of specific proteins, including the immune checkpoint factors PDCD and CD274, and cellular pathways in the leukocytes of the Klf1(K74R) that are in the directions of anti-cancer and/or anti-aging. This study demonstrates the feasibility of developing a transferable hematopoietic/blood system for long-term anti-cancer and, potentially, for anti-aging.
Introduction
Aging of animals, including humans, is accompanied by lifespan-dependent organ deterioration and the occurrence of chronic diseases such as cancer, diabetes, cardiovascular failure, and neurodegeneration (Aman et al., 2021; López-Otín et al., 2013). To extend healthspan and lifespan, various biomedical- and biotechnology-related strategies have been intensively developed and applied, including the therapy of different diseases such as cancer (Bashor et al., 2022; Fontana and Partridge, 2015; Longo and Anderson, 2022). The hematopoietic/blood system is an important biomedical target for anti-aging and anti-cancer research development. Multiple blood cell lineages arise from hematopoietic stem cells (HSCs), with the lymphoid lineage giving rise to T, B, and natural killer (NK) cell populations, whereas the myeloid lineage differentiates into megakaryocytes, erythrocytes, granulocytes, monocytes, and macrophages (Mitchell et al., 2022; Pálovics et al., 2022; Seita and Weissman, 2010). The genetic constituents and homeostasis of the hematopoietic system are regulated epigenetically and via environmental factors to maintain animal health (Mitchell et al., 2022; Montazersaheb et al., 2022).
EKLF, also named KLF1, is a Krüppel-like factor that is expressed in a range of blood cells, including erythrocytes, megakaryocytes, T cells, NK cells, as well as in various hematopoietic progenitors, including common myeloid progenitor , megakaryocyte erythroid progenitor (MEP), and granulocyte macrophage progenitor (Frontelo et al., 2007; Nishizawa et al., 2016; Teruya et al., 2018). The factor regulates erythropoiesis (Perkins et al., 2016) and the differentiation of MEP to megakaryocytes and erythrocytes (Frontelo et al., 2007; Neuwirtova et al., 2013) as well as of monocytes to macrophages (Luo et al., 2004). KLF1 is also expressed in HSC and regulates their differentiation (Hung et al., 2020). The factor can positively or negatively regulate transcription, including the adult β globin genes, through binding of its canonical zinc finger domain to the CACCC motif of the regulatory regions of a diverse array of genes (Hung et al., 2020; Siatecka and Bieker, 2011; Pilon et al., 2011; Tallack et al., 2012). In fact, the zinc finger of KLF1 could recognize/bind to genomic sites with the consensus sequence CCNCNCCC, and recruit different co-activator or co-repressor chromatin remodeling complexes (Neuwirtova et al., 2013; Siatecka and Bieker, 2011).
KLF1 could be sumoylated in vitro and in vivo, and sumoylation of the lysine at codon 74 of mouse KLF1 altered the transcriptional regulatory function (Frontelo et al., 2007) as well as nuclear import (Shyu et al., 2014) of the factor. Surprisingly, homozygosity of a single amino acid substitution, lysine(K) to arginine(R), at the sumoylation site of KLF1 results in the generation of a novel mouse model with healthy longevity. These mice, termed Klf1K74R/K74R or Klf1(K74R), exhibited extended healthspan and lifespan. In particular, the Klf1(K74R) mice showed delay of the age-dependent decline of physical performance, such as the motor function and spatial learning/memory capability, and deterioration of the structure/function of tissues, including the heart, liver, and kidney. Furthermore, the Klf1(K74R) mice appeared to have significantly higher anti-cancer capability than the WT mice (Shyu et al., 2022).
As described in the following, we have since characterized the high anti-cancer capability of the Klf1(K74R) mice with respect to its dependence on the age, gender, and genetic background. More importantly, we have demonstrated that the high anti-cancer ability of these genetically engineered mice could be transferred to wild-type mice (WT) through hematopoietic transplantation of the bone marrow mononuclear cells (BMMNCs). Furthermore, we show that the higher anti-cancer capability and extended lifespan of Klf1(K74R) mice are associated with changes in the global protein expression profile and specific aging-/cancer-associated cellular signaling pathways in their white blood cells (WBCs) or leukocytes.
Results
Characterization of the cancer resistance of Klf1(K74R) mice in relation to age, gender, and genetic background
The Klf1(K74R) mice appeared to be cancer resistant to carcinogenesis as manifested by their lower spontaneous cancer incidence (12.5%) in life than WT mice (75%). The Klf1(K74R) mutation also protected the mice from metastasis of melanoma cells and reduced melanoma growth in the subcutaneous cancer cell inoculation assay (Shyu et al., 2022). Here, we used the pulmonary melanoma foci assay, which has been commonly used for quick analysis of early metastasis (Vaseghi et al., 2023), to further characterize the higher cancer resistance of the Klf1(K74R) mice with respect to the effects of gender/age/genetic background of the mice and the requirement for the homozygosity of the K74R mutation. The cultured melanoma B16-F10 cells used in this assay are poorly immunogenic and highly aggressive, thus being useful for monitoring early metastasis in various immunotherapy studies (Vaseghi et al., 2023; Díaz-García et al., 2018).
It appeared that male as well as female Klf1(K74R) mice in the B6 genetic background had significantly fewer pulmonary melanoma foci than the corresponding WT mice (Figure 1). Because of this result, we used male mice for all of the studies described below. First, both young (2-month-old) and aged (24-month-old) Klf1(K74R) mice had higher anti-metastasis ability against the injected melanoma cells than WT mice of age-dependent groups (Figure 1A and B). Secondly, homozygosity of the K74R substitution was required for the higher cancer resistance of the Klf1(K74R) mice (Figure 1—figure supplement 1). Importantly, the Klf1(K74R) mice in the FVB background also exhibited a high cancer resistance than FVB WT mice by this assay (Figure 1C), suggesting that cancer resistance of Klf1(K74R) mice conferred by the homozygous K74R substitution was likely genetic background-independent. Finally, the higher anti-cancer capability of the Klf1(K74R) mice did not appear to depend on the arginine at codon 74 since Klf1(K74A) mice carrying K→A amino acid substitution at the K74 sumoylation site also exhibited higher anti-metastasis capability than WT mice in the pulmonary foci assay (Figure 1D).
Figure 1 with 1 supplement see all
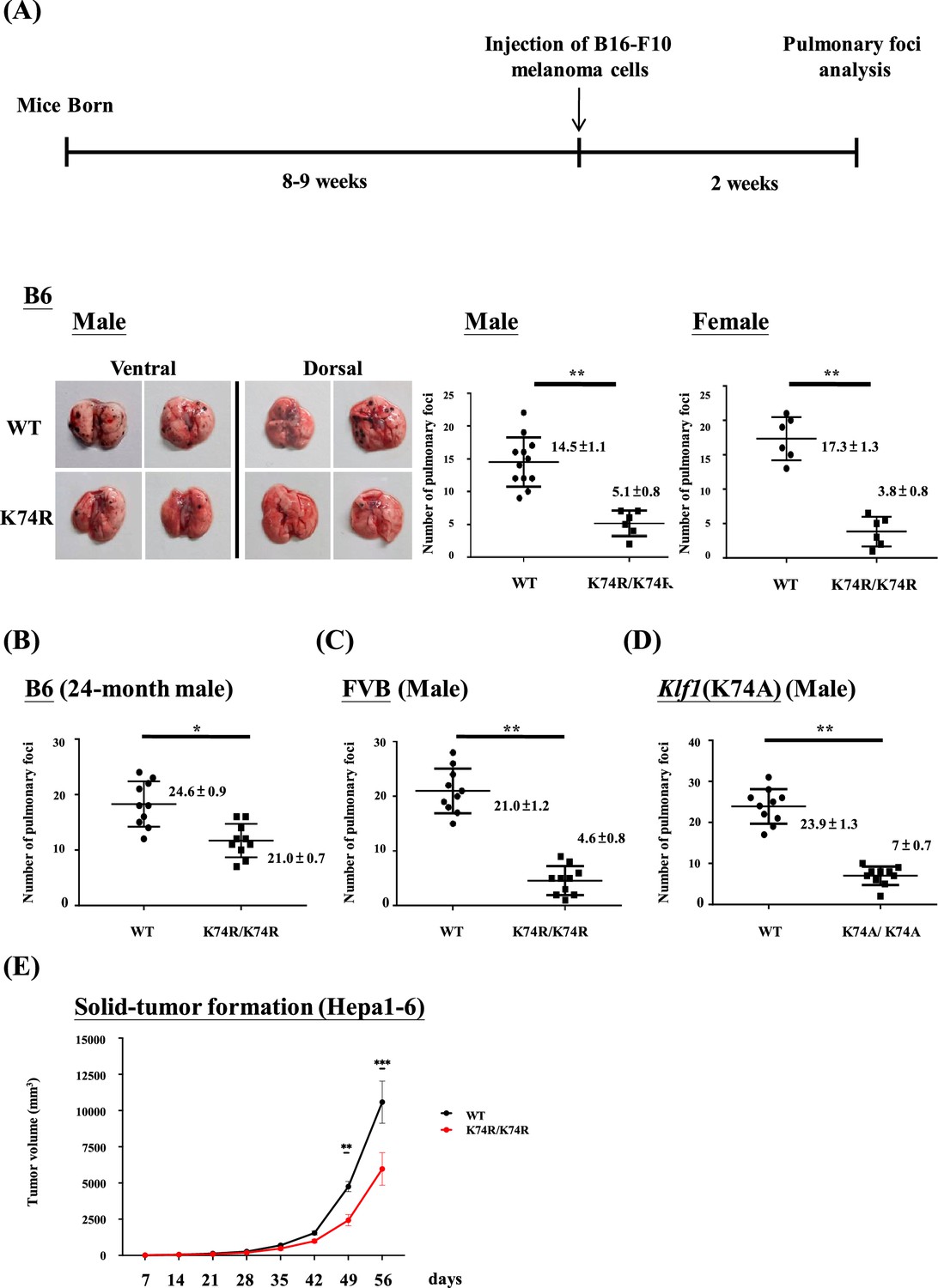
Anti-cancer capability of Klf1(K74R) mice as analyzed by the experimental melanoma metastasis assay.
(A) Flow chart illustrating the strategy of the pulmonary tumor foci assay. Left panels: representative photographs of pulmonary metastatic foci on the lungs of wild-type (WT) and Klf1(K74R) male mice in the B6 background 2 wk after intravenous injection of B16-F10 cells (105 cells/ mouse). Statistical comparison of the numbers of pulmonary foci is shown in the two histograms on the right. N = 10 (male) and N = 7 (female), **p<0.01. Note that only the number of large pulmonary foci (>1 mm diameter) was scored. N > 6, **p<0.01. (B) Pulmonary tumor foci assay of 24-month-old WT and Klf1(K74R) male mice. Statistical comparison is shown in the two histograms. N = 10 (male), *p<0.05. (C) Pulmonary tumor foci assay of male mice in the FVB background. Statistical comparison is shown in the histograph on the right. N = 10, **p<0.01. (D) Pulmonary tumor foci assay of Klf1(K74A) male mice. Statistical comparison of the 3-month-old WT and Klf1(K74A) mice numbers of pulmonary foci is shown in the two histograms. N = 10 (male), **p<0.01. (E) The Klf1(K74R) mice and WT mice were subcutaneously injected with Hepa1-6 cells to form tumors. The tumor volumes were measured once a week using the formula: length × weight × height × π/6. The curves of tumor growth started to show difference between the Klf1(K74R) and WT mice at 49 d after Hepa1-6 cell injection. N > 6, *p<0.05, **p<0.01, and ***p<0.001.
The anti-cancer capability of Klf1(K74R) mice was not restricted to melanoma. As shown by the subcutaneous inoculation assay of Hepa1-6 cells (Figure 1E), Klf1(K74R) mice also carry a higher anti-cancer capability against hepatocellular carcinoma than the WT mice. It thus appears that the Klf1(K74R) mice are resistant to the carcinogenesis of a range of different cancers.
Transfer of the anti-cancer capability and extended lifespan of Klf1(K74R) mice to WT mice via BMT
Since KLF1 is a hematopoietic transcription factor expressed not only in mature blood cells but also in HSCs and hematopoietic stem progenitor cells, this cancer resistance may be transferable by means of BMT. This possibility was tested by using male mice and a standard BMT protocol (Imado et al., 2004). BMMNCs were purified from the bone marrow of 2-month-old CD45.2 Klf1(K74R) or WT mice and injected into the tail vein of CD45.1 WT recipient mice. Blood replacement of recipient mice with 10 Gy γ-irradiation by that of the donor mice reached 90% in the seventh week (Figure 2A and B). After 2 wk, the recipient mice were injected with B16-F10 cells and then sacrificed a further 2 wk later to quantify pulmonary tumor foci. We found that WT mice transplanted with WT BMMNC had similarly high numbers of tumor foci relative to WT mice without BMT (Figures 1A and 2C). However, similar to Klf1(K74R) mice challenged with B16-F10 cells (Figure 1B), WT mice that received BMMNC from Klf1(K74R) mice presented significantly fewer tumor foci on their lungs (Figure 2C). Notably, BMT using 24-month-old donor mice gave similar results, although the effect is relatively small (Figure 2—figure supplement 1A).
Figure 2 with 1 supplement see all
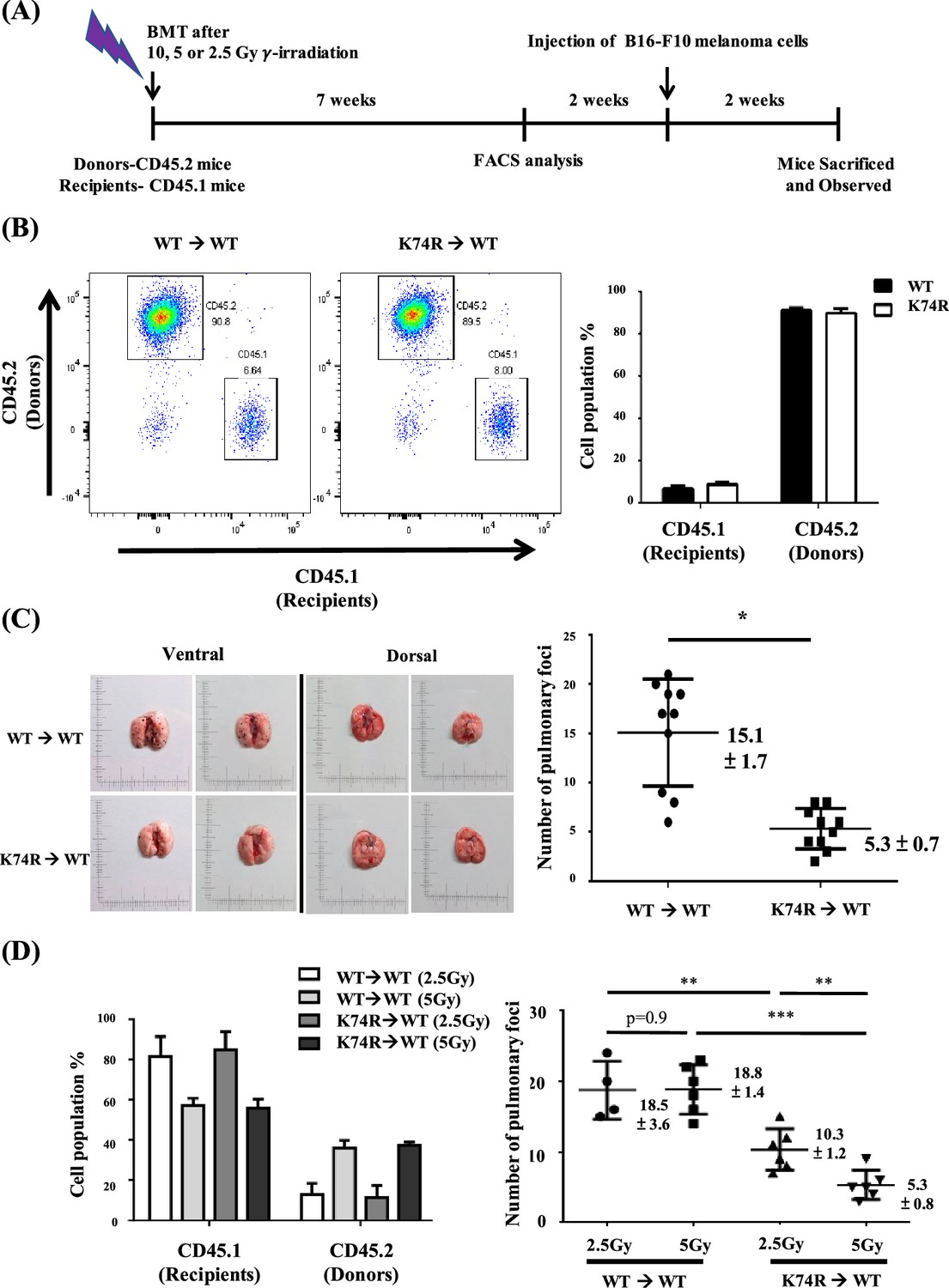
Transfer of cancer resistance of Klf1(K74R) mice to wild-type (WT) mice by bone marrow transplantation (BMT).
(A) Flow chart illustrating the experimental strategy. (B) Fluorescence-activated cell sorting (FACS) analysis of the efficiency of BMT by using 10 Gy γ-irradiation. The percentages of CD45.1/CD45.2 cells in the peripheral blood (PB) of the recipient male mice were analyzed by flow cytometry, with the representative FACS charts shown on the left and the statistical histobar diagram on the right. (C) Transfer of the anti-metastasis capability of 8-week-old Klf1(K74R) male mice to age-equivalent WT male mice by BMT by using 10 Gy γ-irradiation. Left panels: representative photographs of lungs with pulmonary metastatic foci in the recipient WT (CD45.1) mice after BMT from WT (CD45.2) or Klf1(K74R) (CD45.2) donor mice and challenged with B16-F10 cells. Statistical analysis of the number of pulmonary B16-F10 metastatic foci on the lungs is shown in the right histogram. n = 10, *p<0.05. (D) Transplantation of 8-week-old male WT (CD45.1) mice with BMMNC from age-equivalent WT (CD45.2) male mice or from Klf1(K74R) (CD45.2) male mice by using the γ-irradiation dosage 2.5 Gy or 5 Gy. The histobar diagram comparing the percentages of CD45.1 and CD45.2 PB cells of the recipient WT mice after BMT is shown on the left. The statistical analysis of the average number of pulmonary foci on the lungs of recipient WT mice after BMT and injected with the B16-F10 cells is shown in the right histogram, N = 6. **p<0.01, ***p<0.001.
In order to determine whether WT mice having more restricted blood replacement upon BMT from Klf1(K74R) mice still exhibited a higher anti-cancer capability, we also carried out BMT experiments with lower doses of γ-irradiation. BMT using two lower doses of γ-irradiation (2.5 Gy/5 Gy) still resulted in the transfer of cancer resistance from Klf1(K74R) to WT mice. Approximately 40% of recipient blood cells were substituted by donor cells upon BMT with 5 Gy γ-irradiation. Consequently, at that irradiation dosage, BMT from Klf1(K74R) mouse donors reduced the average number of pulmonary tumor foci in recipient WT mice to 5. On the other hand, only 20% blood replacement was achieved in the recipient mice with 2.5 Gy γ-irradiation (Figure 2D). However, the WT mice receiving BMT from Klf1(K74R) mice again developed less number (~10/mouse) of pulmonary tumor foci than those WT mice receiving BMT from the WT mice (Figure 2D). Thus, even at a low level of 20% blood replacement, BMT still enabled effective transfer of cancer resistance from Klf1(K74R) mice to WT mice.
In addition, we also attempted to transfer the extended lifespan characteristics of the Klf1(K74R) mice to WT mice by BMT. Significantly, the median lifespan of WT mice receiving BMT from Klf1(K74R) mice was 5 mo longer than that of WT mice receiving BMT from WT mice (Figure 2—figure supplement 1B). Thus, the longer lifespan characteristics of the Klf1(K74R) mice were also transferable via BMT.
Inhibition of tumor growth by transplanted BMMNC from Klf1(K74R) mice
Our experiments indicated that Klf1(K74R) bone marrow carried the anti-metastasis capability that prevented melanoma cell colonization in the lungs of recipient mice (Figures 1 and 2). To determine whether Klf1(K74R) BMT could inhibit tumor growth, we examined the effects of BMT on growth of tumors with B16-F10-luc cells. As outlined in Figure 3A, 10 d after injection of cancerous cells, the formation of bioluminescent signals in the recipient mice was confirmed by the observation of in vivo bioluminescence. The following day, we transplanted the recipient mice with BMMNC from WT or Klf1(K74R) mice and then measured the intensities of bioluminescence signals from tumors 7 and 14 d later. As shown, tumor growth in mice subjected to BMT from Klf1(K74R) mice was significantly slower relative to those receiving BMMNC from WT mice (Figure 3B and C). Thus, Klf1(K74R) BMMNC indeed can inhibit the growth of tumors more effectively than WT BMMNC.
Figure 3
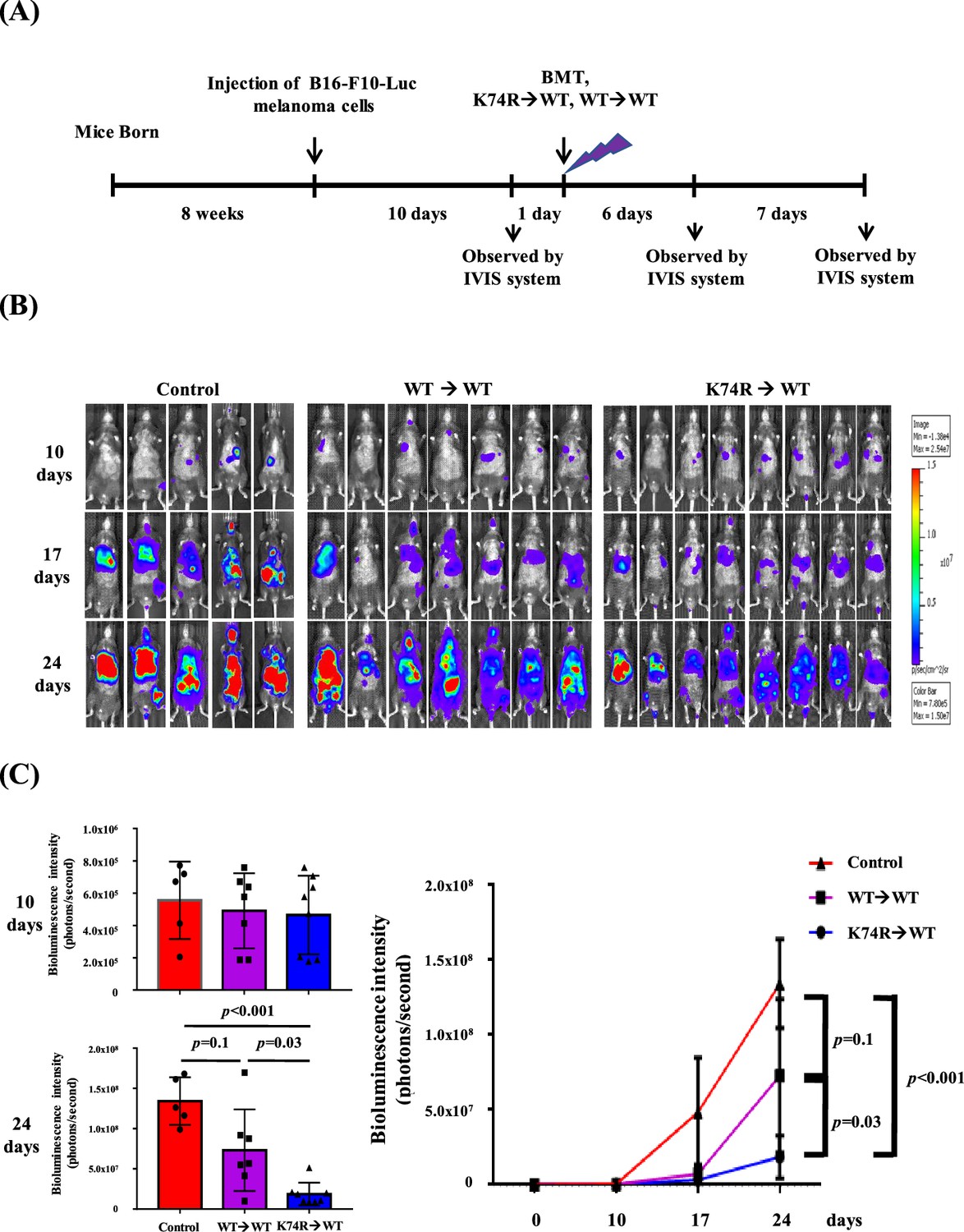
Inhibition of tumor growth in wild-type (WT) mice by bone marrow transplantation (BMT) from Klf1(K74R) mice.
(A) A flow chart of the experiments. Luciferase-positive B16-F10 cells were injected into the tail vein of 8-week-old WT male mice (day 0). The mice were then transplanted with bone marrow mononuclear cells (BMMNCs) from WT or Klf1(K74R) male mice on day 11 after the luciferase-positive B16-F10 cell injection. In vivo imaging system (IVIS) was used to follow the tumor growth in mice on days 0, 10, 17, and 24, respectively. (B) Representative images of bioluminescence reflecting the luciferase activity from melanoma cancer cells in mice. The color bar indicates the scale of the bioluminescence intensity. (C) Statistical analysis of the intensities of bioluminescence in the cancer-bearing mice (WT→WT, purple, N = 7; Klf1(K74R)→WT, blue, N = 8; control [no BMT], red, N = 3).
Differential expression of specific immune-, aging-, and/or cancer- associated biomolecules in the blood of Klf1(K74R) mice
In general, the cell populations in peripheral blood (PB) were not affected much by the K74R substitution, as shown by CBC analysis (Shyu et al., 2022), although flow cytometry analysis of PB from WT and Klf1(K74R) mice of different ages showed that the population of NK(K74R) cells in 24-month-old Klf1 (K74R) mice was higher than the WTNK cells in 24-month-old WT mice (Figure 4—figure supplement 1). In parallel, the K74R substitution did not affect much the expression of Klf1 gene in the different types of hematopoietic cells. As shown in Figure 4—figure supplement 2, the K74R substitution did not alter the expression levels of KLF1 protein in leukocytes from PB (Figure 4—figure supplement 2A, left panels), although the Klf1 mRNA level of Klf1 (K74R) leukocytes was 10–20% lower than that of WT leukocytes (Figure 4—figure supplement 2A, right panel). Previous studies and databases have shown that Klf1 mRNA and KLF1 protein are expressed in the erythroid lineage (Frontelo et al., 2007) and megakaryocytes (Frontelo et al., 2007; Neuwirtova et al., 2013), and the Klf1 mRNA is expressed in HSC (Hung et al., 2020) (bio-GPS database), NK cells (bio-GPS database), naïve T cells (Teruya et al., 2018), and Treg cells (Teruya et al., 2018). By western blotting (WB) analysis (Figure 4—figure supplement 2B) and RT-qPCR analysis (Figure 4—figure supplement 2C), we found that the Klf1 gene is expressed in B220+ B cells, CD3+ T cells, and NK cells of both WT and Klf1(K74R) mice, but the levels are much lower than that of the erythroid MEL cells.
We first used RT-qPCR to analyze the levels in PB cells of mRNAs encoding the immune checkpoint genes (ICGs) PDCD/CD274 (Iwai et al., 2017) in view of the cancer resistance of Klf1(K74R) mice (Figure 4) as well as increased levels of PDCD and CD274 in aged or tumorigenic mice (Jiang et al., 2019). As shown in Figure 4, the mRNA levels of Pdcd and Cd274 in the PB, B cells, and T cells of WT mice were both increased during aging. In contrast, the mRNA levels and protein levels of these two genes were lower in 3-month-old Klf1(K74R) mice than the age-matched WT mice, and they remained low during aging of the Klf1(K74R) mice. Significantly, RNAi knockdown of Klf1 expression reduced Pdcd and Cd274 mRNA levels in splenic CD3+ T cells (Figure 4C). In an interesting correlation, overexpression of KLF1 would increase the expression of Cd274 in mouse CD4+ T cells (Teruya et al., 2018). These data together indicate that KLF1 positively regulates the transcription of the Pdcd and Cd274 genes, directly or indirectly, and likely the transcriptomes of a diverse range of hematopoietic cells. Since the levels of KLF1 protein in CD3+ T (K74R) cells and WT CD3+ T cells were similar (Figure 4—figure supplement 2B), change of the expression of Pdcd and Cd274 genes (Figure 4) was most likely caused by the loss of the sumoylation of KLF1(K74R), which would alter its transactivation capability (Siatecka and Bieker, 2011). Interestingly, Pdcd appeared to be a direct regulatory target of KLF1. As shown by ChIP-qPCR analysis, KLF1 was bound at the CACCC box (Box3) at –103 of the Pdcd promoter in CD3+ T cells from both WT and Klf1(K74R) mice (Figure 4—figure supplement 3). The details of the role of sumoylation in the transcriptional activation of Pdcd by KLF1 await future investigation. As expected, lower levels of Pdcd and Pdcd1 expression were also observed in the PB of mice receiving BMT from Klf1(K74R) mice (Figure 4—figure supplement 2D). These findings together suggest that the low tumorigenesis rate of Klf1(K74R) mice arises in part from the low expression of the two ICGs, Pdcd and Cd274, as a result of K74R substitution in the KLF1 protein.
Figure 4 with 4 supplements see all
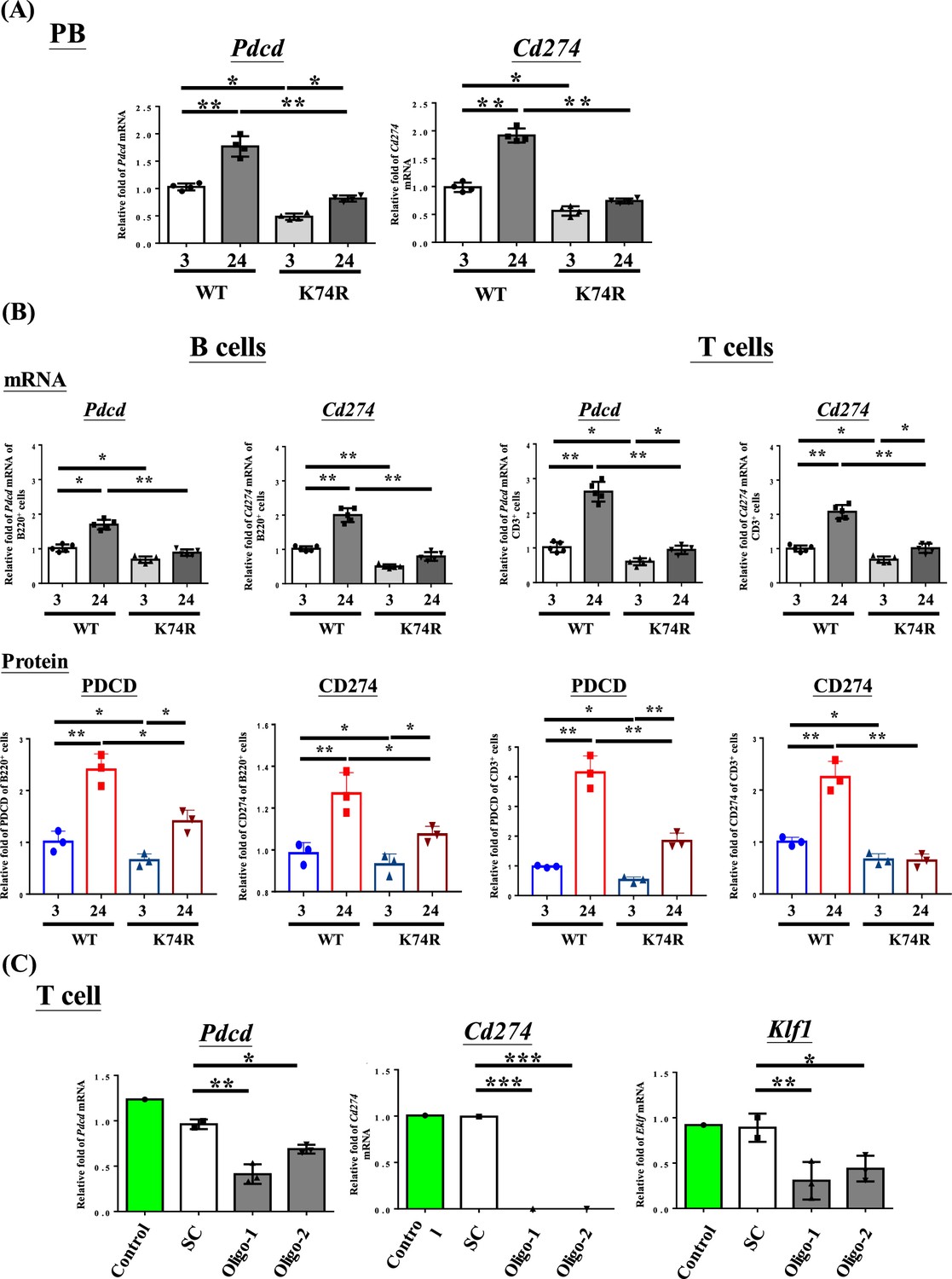
Decrease in Pdcd and Cd274 expression in blood cells of Klf1(K74R) mice.
(A) Levels of Pdcd and Cd274 mRNAs in the peripheral blood (PB) of wild-type (WT) and Klf1(K74R) male mice at the ages of 3 mo and 24 mo, respectively, as analyzed by RT-qPCR. Note the relatively low levels of Pdcd and Cd274 mRNAs in the Klf1(K74R) mice at both ages in comparison to the WT mice. (B) Upper panels: comparison of the mRNA levels of Pdcd and Cd274 of CD3+ T cells and B220+ B cells isolated from the PB of 8-week-old WT and Klf1(K74R) male mice. N = 5. *p<0.05; **p<0.01. Lower panels: comparison of the protein levels of PDCD and CD274, as analyzed by flow cytometry, of CD3+ T cells and B220+ B cells from 8-week-old WT and Klf1(K74R) male mice. N = 3. *p<0.05; **p<0.01. (C) Comparison of the levels of Pdcd, Cd274, and Klf1 mRNAs, as analyzed by RT-qPCR, in CD3+ T cells, which were isolated from splenocytes, without or with RNAi knockdown of Klf1 mRNA. Two oligos (oligo-1 and oligo-2) were used to knock down Klf1 mRNA by ~60–70%, which resulted in the reduction of Pdcd mRNA level by 30–60% and nearly complete depletion of Cd274 mRNA. Control, T cells transfected with GFP-plasmid; SC, T cells transfected with scrambled oligos. N > 3. *p<0.05; **p<0.01; ***p<0.001.
We also examined, by bead-based multiplex assay (Aira et al., 2019; Menees et al., 2021), the expression patterns of several aging- and/or cancer-associated cytokines. As shown in Figure 4—figure supplement 4, there was no significant difference in the serum levels of IL-1β, IL-2, IL-10, IL-12p70, INF-γ, or TNF-α between WT and Klf1(K74R) mice at 24-month-old. In contrast, the level of IL-4, an anti-inflammatory cytokine (Ul-Haq et al., 2016) beneficial to the hippocampus of aging mice (Boccardi et al., 2018), in 24-month-old Klf1(K74R) mice was three- to fourfold higher than the 24-month-old WT mice. On the other hand, the level of IL-6, a key factor in chronic inflammatory diseases, autoimmunity, cytokine storm, and cancer (Menees et al., 2021; Rea et al., 2018), increased only moderately during aging of the Klf1(K74R) mice (Figure 4—figure supplement 4). Thus, similar to PDCD and CD274, the altered expression of some of the cytokines in the blood likely also contributes to the anti-aging and/or anti-cancer characteristics of the Klf1(K74R) mice.
Comparative proteomics analysis of the leukocytes of Klf1(K74R) mice and WT mice
We proceeded to examine age-dependent cell-intrinsic changes in the proteomes of the leukocytes from the WT and Klf1(K74R) mice in two different age groups. A total of 259 and 306 differentially expressed proteins (DEPs) were identified between the two age groups for the WT and Klf1(K74R) mice, respectively (Figure 5A). To understand the correlations of these proteins with aging and cancer, we performed the Gene Set Enrichment Analysis (GSEA) and found that the age-dependent DEPs changed in the concordant direction in the WT and Klf1(K74R) mice were enriched for several known aging-related pathways, for example, IL-6-JAK-STAT3 signaling, DNA repair, etc. (Fabian et al., 2021; Figure 5B). Meanwhile, the age-dependent DEPs changed in the reverse directions in the WT and Klf1(K74R) mice were enriched for nine other aging-related pathways (Figure 5C).
Figure 5
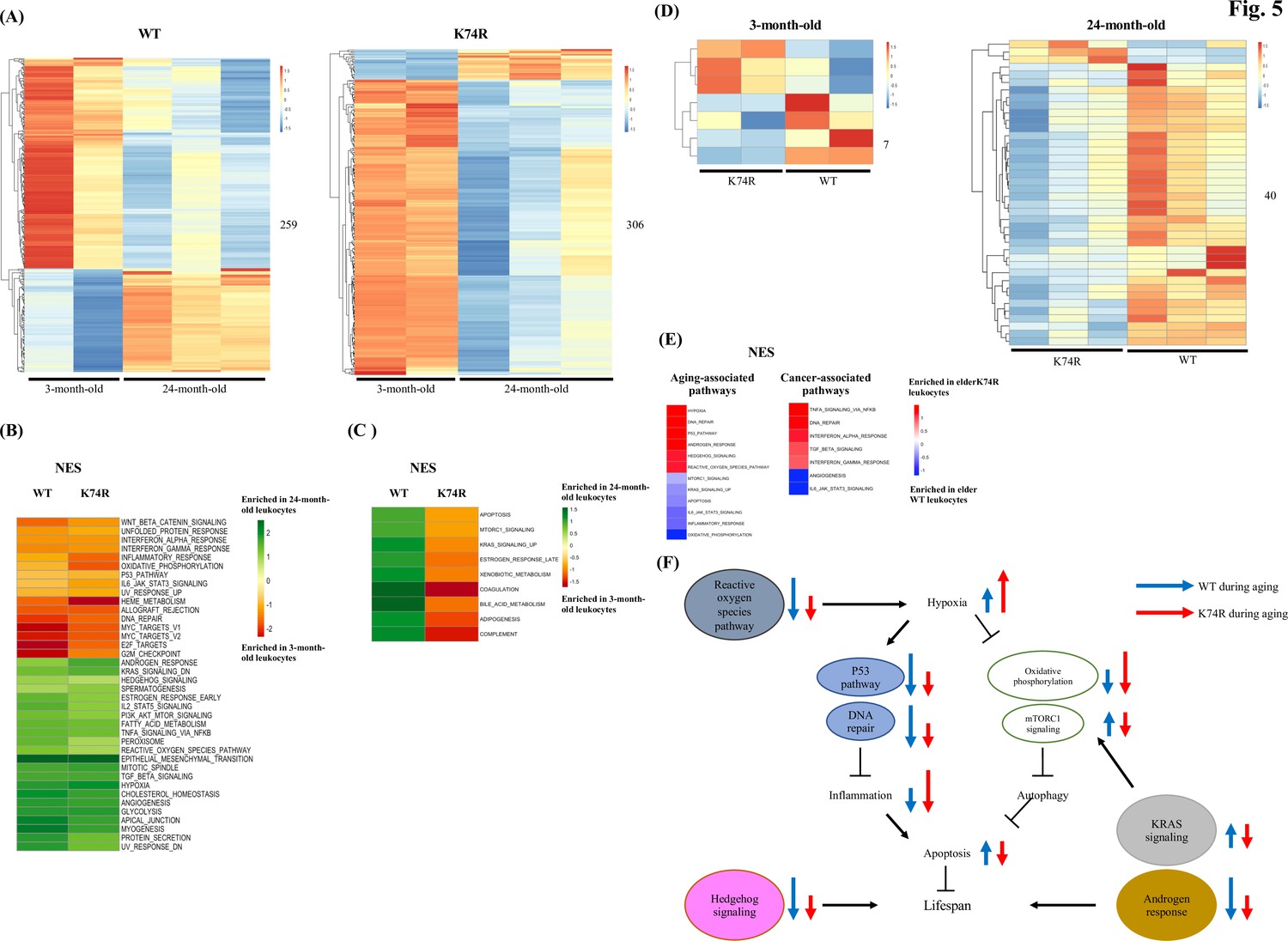
Proteomics analysis of mouse leukocytes.
(A) Heatmap plots representing age-dependent differentially expressed proteins (DEPs) in the leukocytes from the wild-type (WT) male mice (left) and Klf1(K74R) male mice (right). Differential test threshold: expression fold change >1.5 and p-value <0.01. (B, C) Pathway analysis of the age-dependent DEPs changed in the concordant (B) and reverse (C) directions, respectively, in the WT and Klf1(K74R) mice. NES, normalized enrichment score. (D) Heatmap plots representing strain-dependent DEPs in leukocytes of 3-month-old (left) and 24-month-old (right) mice. (E) Pathway analyses of the strain-dependent DEPs in elder WT leukocytes (enrichment indicated by blue color) and elder Klf1(K74R) leukocytes (enrichment indicated by red color). The aging- and cancer-associated pathways are presented in the left and right diagrams, respectively. (F) A model of the regulation of the mouse lifespan by different cellular pathways in the leukocytes. The inter-relationship of 11 aging-associated cellular pathways differentially expressed in the leukocytes of Klf1(K74R) male mice in comparison to the WT male mice, among themselves and with respect to the regulation of lifespan (for references, see text), is depicted here. The directions and lengths of the arrows indicate changes in the individual pathways, upregulation (up arrows) or repression (down arrows), in the leukocytes during aging of the WT male mice (blue arrows) and Klf1(K74R) male mice (red arrows), respectively.
We further performed DEP analyses between WT and Klf1(K74R) mice and identified strain-dependent DEPs in the two age groups. As shown in Figure 5D, only 7 DEPs were identified in the 3-month-old mice but 40 DEPs in the 24-month-old ones. Of the 40 DEPs in the elder mice, 3 and 37 were upregulated in Klf1(K74R) and WT mice, respectively (Figure 5D). Significantly, GSEA of these DEPs showed that elder Klf1(K74R) leukocytes were enriched for the anti-aging pathways related to hypoxia, p53 signaling, etc. (Yeo, 2019; Zhu et al., 2021), while the elder WT leukocytes were enriched for the aging-associated pathways related to apoptosis, mTORC1 signaling, etc (Yeo, 2019; Hamarsheh et al., 2020). On the other hand, the DEPs in elder Klf1(K74R) leukocytes were also enriched for anti-cancer pathways related to the IFN-α response, TGF-β signaling, etc. (Ni and Lu, 2018; Principe et al., 2014; Zitvogel et al., 2015; Figure 5E), while DEPs in the elder WT leukocytes were enriched for the pro-cancer pathways related to IL-6-JAK-STAT3 signaling and angiogenesis (Zhao et al., 2016). These data together have demonstrated that Klf1(K74R) leukocytes contribute to their anti-cancer capability and long lifespan through several specific cellular signaling pathways.
Higher in vitro cancer cell cytotoxicity of the NK(K74R) cells than WT NK cells
The higher anti-cancer capability of the Klf1(K74R) mice could result from combined contributions by different types of the hematopoietic cells, for example, the NK(K74R) cells. NK cells are effector lymphocytes of the innate immune system that can rapidly destroy tumor cells and against infectious pathogens (Roncagalli et al., 2005). Also, NK cells do not require pre-stimulation for the treatment of cancer cells (Yu, 2023). Furthermore, the NK lineage could be fully reconstituted 2 wk after BMT (Ferreira et al., 2019). We tested the in vitro cancer cell cytotoxicity ability of NK(K74R) cells from 3-month-old Klf1(K74R) mice in comparison to the WT NK cells. As shown in Figure 6, NK(K74R) cells exhibited significantly higher cytotoxicity against the B16-F10 melanoma cells as well as Hepa1-6 hepatocellular carcinoma cells than WT NK cells. This data, together with the higher level of NK cells in aged Klf1(K74R) mice (Figure 4—figure supplement 1), indicated that in parallel to the leukocytes (Shyu et al., 2022), NK cells also play an important role in the higher anti-cancer capability of the hematopoietic blood system of Klf1(K74R) mice, as exemplified in Figures 2 and 3.
Figure 6

Higher in vitro cancer cell cytotoxicity of NK(K74R) cells than wild-type (WT) natural killer (NK) cells.
B16-F10 and Hepa1-6 cells were labeled with Calcein-AM (BioLegend) and co-cultured with NK cells in different E:T ratio for 4 hr. Killing rates of B16-F10 and Hepa 1–6 cancer cells by NK cells were determined by the intensities of fluorescence in the medium in comparison to the controls of spontaneous release and maximal release.
Discussion
Because of the complexity and intercrosses of the different pathways regulating the health and the aging process, genetic manipulation of non-human animals (Bin-Jumah et al., 2022; Hofmann et al., 2015), and non-genetic approaches on animals including humans (Fontana and Partridge, 2015; Ocampo et al., 2016; Varady et al., 2022), targeting these pathways inevitably leads to moderate-to-severe side effects such as body weight loss and adiposity. With respect to the above, the Klf1(K74R) mice (Shyu et al., 2022) are ideal as an animal model for further insightful understanding of the aging process as well as for biomedical development of new anti-aging tools and approaches. In the current study, we demonstrate that the healthy longevity of the Klf1(K74R) mice, as exemplified by their higher tumor resistance, is likely to be gender/genetic background independent and irrespective of the amino acid change at K74 of KLF1. Furthermore, we show that the Klf1(K74R) mice are resistant to carcinogenesis of not only melanoma, but also other cancers such as hepatocellular carcinoma. This correlates well with the higher cytotoxicity in vitro of NK(K74R) cells than WT NK cells against the same two types of cancer cells. Significantly, we also show that one-time transplantation of the BMMNC(BMT) could confer the recipient WT mice extended lifespan and higher tumor-resistance capability, the latter of which could be achieved by 20% of substitution of the blood of the recipient WT mice with that from the Klf1(K74R) mice. These findings together have interesting implications in the development of this novel hematopoietic blood system for future medical application in anti-disease and anti-aging.
The anti-cancer capability of the Klf1(K74R) mice has rendered them relatively free from spontaneous cancer occurrence (Shyu et al., 2022), which is also reflected by their resistance to tumorigenesis of the B16-F10 cells and Hepa1-6 in the anti-cancer assays (Figures 1—3). Furthermore, the cancer resistance of Klf1(K74R) mice appears to be independent of the gender, age, and genetic background (Figure 1). The anti-metastasis property of the Klf1(K74A) mice in the pulmonary foci assay (Figure 1D) also indicates that the anti-cancer capability of the Klf1(K74R) mice is not due to the structural and/or post-translational properties of the arginine introduced at codon 74 of KLF1. Importantly, we have shown that the anti-cancer capability and the extended lifespan characteristics of Klf1(K74R) mice are transferrable through BMT (Figures 2 and 3, Figure 2—figure supplement 1A). In particular, we show that BMMNC from Klf1(K74R) mice (Figure 2, Figure 2—figure supplement 1A) could confer 2-month-old WT recipient mice with the anti-cancer capability against melanoma. Furthermore, ~20% of blood substitution would allow the recipient mice to become cancer-resistant in the pulmonary foci assay (Figure 2D). It is likely that homozygosity, but not heterozygosity, of K74R substitution causes one or more types of the hematopoietic cells to gain new functions, such as the higher cancer cell-killing capability of NK(K74R) cells (Figure 6).The data of Figure 2D further suggest that the amount of specific types of cells such as NK(K74R) in the 20% of blood cells carrying homozygous Klf1(K74R) alleles in the recipient mice upon BMT is sufficient to confer the mice a higher anti-cancer ability.
HSC therapy for different diseases (Hifumi et al., 2017; Morgan et al., 2017; Giannaccare et al., 2020; Wu et al., 2020) including cancer has been intensively explored and practiced such as leukemia and neuroblastoma (Daver et al., 2020; Mora, 2022). Also, certain characteristics of the young mice could be transferred to old mice via heterochronic parabiosis or heterochronic transplantation (Conboy et al., 2005; Das et al., 2019; Goodell and Rando, 2015; Kang et al., 2020). Similarly, plasma proteins from human umbilical cord blood can revitalize hippocampal function and neuroplasticity in aged mice (Castellano et al., 2017; Mehdipour et al., 2020). Previously, we have shown that multiple injections of LSK-HSC(K74R) from 3-month-old Klf1(K74R) mice to 25-month-old WT mice would extend the lifespan of the latter by about 4 months (Shyu et al., 2014). In the current study, we further show that 2-month-old WT mice receiving one-time transplantation of BMMNCs from 2-month-old Klf1(K74R) mice would live 5.5 mo longer than the controls (Figure 2—figure supplement 1B). Thus, transplantation/transfer of the blood MNC carrying homozygous mutation at the sumoylation site of KLF1 could be developed as a new approach for long-term anti-aging, anti-cancer, and likely rejuvenation.
The tumorigenesis resistance and long lifespan exhibited by the Klf1(K74R) mice are most likely due to changes in the transcription regulatory properties of the mutant KLF1(K74R) protein relative to WT (Siatecka et al., 2007). As exemplified in Figure 4A and B, expression levels of the ICGs Pdcd and Cd274 in the PB, B, and T cells of Klf1(K74R) mice are reduced in comparison to the WT mice. Notably, cancer incidence increases with aging (Aunan et al., 2017), which is accompanied by increased expression of PDCD and CD274 (Lages et al., 2010). The lower expression of ICGs would contribute to the anti-cancer capabilities of the Klf1(K74R) blood to fight against cancer (Figures 1—3) and to extension of the lifespan of cancer-bearing mice (Shyu et al., 2022). Indeed, similar to ICGs, the expression levels of several cytokines in the Klf1(K74R) blood/serum are also different from the WT blood, and some of the changes during aging or carcinogenesis in the Klf1(K74R) blood are opposite to the blood/serum of WT mice (Menees et al., 2021; Figure 4—figure supplement 4).
The transcriptome data have been used to dissect the regulation of leukocyte aging (Fabian et al., 2021; Aira et al., 2019; Schaum et al., 2020). In addition, proteomic analysis has revealed the signaling pathways that regulate aging of specific types of leukocytes such as the lymphocyte and neutrophils cells (Zhou et al., 2014; Zhang et al., 2015). In this study, we have performed proteomics analysis of leukocytes from WT and Klf1(K74R) mice in two age groups and found that for the elder mice the strain-dependent DEPs in the leukocytes are enriched for a number of signaling pathways. Among these signaling pathways, at least 12 of them are closely associated with the aging process, which include hypoxia and DNA repair (Figure 5E). As summarized by the model in Figure 5F, it appears that changes in these pathways in the elder Klf1(K74R) leukocytes relative to the elder WT leukocytes are mostly in the direction of anti-aging. The data of Figure 5 together strongly suggest that the KLF1(K74R) amino acid substitution causes a change in the global gene expression profile of the leukocytes, which contributes to the high anti-cancer capability and long lifespan of the Klf1(K74R) mice.
In summary, we have characterized the cancer resistance of the Klf1(K74R) mice, among their other healthy characteristics, in relation to gender, age, and genetic background. We also have identified cell populations, gene expression profiles, and cellular signaling pathways of the WBCs of young and old mutant mice in comparison to the WT ones, which are changed in the anti-cancer and/or anti-aging directions. Finally, the transferability of the cancer resistance and extended lifespan of the mutant mice via transplantation of BMMNC suggests the possibility of future development of hematopoietic blood cells genome-edited at the conserved sumoylation site of KLF1 for anti-cancer and the extension of healthspan and/or lifespan in animals including humans.
Animals
All the animal procedures were approved by the Institute of Animal Care and Use Committees (IACUC) at Academia Sinica. The IACUC numbers are 17-02-1052, 17-12-1142, 20-10-1528, BioTReC-110-M-005, and BioTReC-111-M-004, respectively.
Materials and methods
Mice
C57BL/6, B6, and FVB mice were purchased from Jackson Laboratories (Bar Harbor, ME). The B6 Klf1(K74R), B6 Klf1(K74A), and FVB Klf1(K74R) mice were established with the assistance of the Transgenic Core Facility, IMB, Academia Sinica, Taiwan. As described previously (Shyu et al., 2022), the K74R mutation was introduced by homologous recombination into exon 2 (E2) of the Klf1 gene of B6 mice by means of a recombinant retrovirus containing the construct loxP-PGK-gb2-neo-loxP-E2 (K74R), before excising the neomycin (neo) selection marker by crossing with Ella-Cre mice. The heterozygous Klf1(K74R/+) mice were then crossed to obtain homozygous mutant Klf1(K74R/K74R) mice, hereafter termed Klf1(K74R) mice.
On the other hand, Klf1(K74A) mice were generated by using the CRISPR/Cas9 system. Female B6 mice (7–8-week-old) were mated with B6 males and the fertilized embryos were collected from the oviducts. For oligos injection, Cas mRNA (100 ng/μl), sgRNA (50 ng/μl), and donor oligos (100 ng/μl) were mixed and injected into the zygotes at the pronuclei stage. The F0 mice were genotyped by PCR and DNA sequencing. The heterozygous Klf1(K74A/+) mice were crossed to establish the germ-line stable homozygous Klf1(K74A) F1 strain.
Klf1(K74R) mice in the FVB background were generated using an in vitro fertilization strategy. Briefly, sperm from male B6 Klf1(K74R) mice was used to fertilize FVB mouse oocytes. In vitro fertilizations of FVB oocytes were carried out consecutively for five generations. The resulting chimeric mice with >90% FVB background were then crossed with FVB mice for another five generations or more.
Cell lines
Request a detailed protocolMurine B16-F10 melanoma cell line was purchased from ATCC (CRL-6475). B16-F10 cells expressing luciferase (B16-F10-luc) were generated by infection of recombinant lentivirus stably expressing luciferase luc2 under the control of hEF1A-eIF4G promoter (InvivoGen) (Lages et al., 2010). All cell lines were derived from cryopreserved stocks split fewer than three times and were confirmed as mycoplasma-free prior to use. B16-F10 cells were cultured at 37°C and 5% CO2 in Dulbecco's Modified Eagle Medium (DMEM) supplemented with 10% fetal bovine serum (FBS), 1% penicillin/streptomycin, and 2 mM l-glutamine. B16-F10-luc cells were selected at 37°C and 5% CO2 in a DMEM supplemented with 0.2 mg/ml zeocin (Invitrogen), 10% FBS, 1% penicillin/streptomycin, and 2 mM l-glutamine. The hepatocellular carcinoma cell line, Hepa1-6, was obtained from Dr. Da-Liang Ou’s lab at the College of Medicine, National Taiwan University. The line was maintained at 37°C and 5% CO2 in Gibico DMEM, a high glucose commercial medium containing 10% FBS (Gibico) and 1% penicillin-streptomycin.
Pulmonary melanoma foci assay
Request a detailed protocolCultured B16-F10 melanoma cells (1 × 105 cells/mouse) were injected into the tail vein of 8- to 9-week-old or 24-month-old Klf1(K74R) and WT mice with or without bone marrow transplantation (BMT). Two weeks after injection, the mice were sacrificed and the number of tumor foci on their lungs was quantified (Narasimhan et al., 2020).
Subcutaneous cancer cell inoculation assay of Hepa1-6
Request a detailed protocolThe protocol is similar to that used by Han et al., 2017. After anesthetization of the mice with 3% isoflurane, 2 × 106/50 ul of Hepa1-6 cells in serum-free DMEM were mixed with 50 ul of Corning Matrigel Matrix and injected between skin and muscle of the hind limb. The size of the tumors was measured and recorded once a week with Mitutoyo Vernier Caliper. The volume was calculated using the formula: length × weight × height × π/6.
Flow cytometric analysis and cell sorting
Request a detailed protocolSingle-cell suspensions of the PB cells and spleen tissue of B6 mice were prepared by lysing red blood cells and then passing them through a 40-μm cell strainer (Falcon). BMMNCs were prepared as described in the section ‘Bone marrow transplantation’. The PB cells and splenocytes were stained extracellularly for 30 min at room temperature using different combinations of the following antibodies: anti-CD45.1 (eBioscience), anti-CD45.2 (eBioscience), anti-CD3e (eBioscience), anti-CD45R (eBioscience), anti-NK1.1 (eBioscience), anti-NKp46 (eBioscience), anti-Gr-1 (eBioscience), CD11b (eBioscience), anti-PDCD (eBioscience), and anti-CD274 (eBioscience). The various hematopoietic progenitor cell compartments of bone marrow were also stained extracellularly for 30 min at room temperature by using different combinations of the following antibodies: anti-lineage (eBioscience), anti-c-Kit (eBioscience), anti-Sca-1 (eBioscience), anti-CD34 (eBioscience), and anti-Flt-3 (eBioscience). All the immunostained cells were subsequently washed with 1% phosphate buffered saline (PBS) three times and resuspended for fluorescence-activated cell sorting (FACS) analysis and sorting. Small amounts of the cell samples were run on a FACS Analyzer LSRII-12P (BD Bioscience) to determine the proportions of different cell preparations. FACS AriaII SORP (BD Bioscience) was then used to sort the indicated cell populations. The detailed gating subsets for all the cell as described above are shown in Supplementary file 1a. Data analysis was performed using the FlowJo software.
RNAi knockdown of Klf1 mRNA from T cells
Request a detailed protocolCD3+ T cells isolated by sorting were cultured in RPMI 1640 medium for 1 d for recovery and then transfected with EGFP-plasmid (control), scrambled oligonucleotides (SC control), Klf1 knockdown oligonucleotide 1 (oligo 1), or Klf1 knockdown oligonucleotide 2 (oligo 2) in a 96-well plate for 48 hr using a LONZA electroporation kit (P3 Primary Cell 4D-Nucleofector X Kit) and machine (4D-Nucleofector Core Unit). Then, the cells were lysed using a PureLink RNA Mini kit (Life Technologies) and analyzed by RT-qPCR.
RT-qPCR analysis
Request a detailed protocolTotal RNAs of B cells, T cells, and leukocytes/WBCs from the PB or spleen of Klf1(K74R) mice and WT mice, respectively, were extracted using a PureLink RNA Mini kit (Life Technologies). The RNAs were reverse-transcribed by means of oligo-dT primers, Maxima H Minus Reverse Transcriptase (Thermo Scientific), and SYBR Green reagents (Applied Biosystems). RT-qPCR was performed using a LightCycler Nano machine (Roche). Gene-specific primers for Klf1, Pdcd, Cd274, and Gapdh were designed using Vector NTI Advance 9 software according to respective mRNA sequences in the NCBI database (primer sequences are available upon request). The PCR primers for analysis of Klf1 mRNA had been used and verified in a previous study (Hung et al., 2020). Expression levels of mRNAs were normalized to that of endogenous Gapdh mRNA.
ChIP-qPCR
Request a detailed protocolThe ChIP-qPCR analysis followed the procedures by Daftari et al., 1999. Extracts were prepared from formaldehyde cross-linked mouse CD3+ T cells and erythroleukemia cell line (MEL), sonicated, and then immune-precipitated (IP) with anti-KLF1 (Thermo) or purified rabbit IgG. The chromatin DNAs in the precipitates were purified and analyzed by qPCR in the Roche LightCycle real-time system. Sequences of the DNA primers used for qPCR are shown in Supplementary file 1b.
Western blotting
Request a detailed protocolWe adopted a previously described WB procedure (Green and Sambrook, 2012). Leukocytes/WBCs of Klf1(K74R) mice and WT B6 mice, respectively, were collected from RBC lysis buffer-treated PB. CD3+ T cells, B220+ B cells, and NK cells from the spleen were sorted using FACS AriaII SORP (BD Bioscience). The cell pellets were lysed in sample buffer and run on SDS-PAGE gels. WB with anti-KLF1 (Abcam) and anti-actin (Sigma) antibodies was then used to analyze and compare the levels of KLF1 and actin proteins.
In vivo bioluminescence imaging
Request a detailed protocolKlf1(K74R) and WT B6 mice were physically restrained and 1 × 105 B16-F10-luc cells/mouse were intravenously injected into their tail vein. Ten days after melanoma cell inoculation, mice were anesthetized for 5 min and injected intraperitoneally with D-luciferin (300 mg/kg of body weight). Fifteen minutes after maximum luciferin uptake, the mice were subjected to imaging of the lung and liver regions in an IVIS 50 Bioluminescence imager (Caliper Life Sciences) to determine metastatic burden. The same mice were used the next day as recipients of BMT from donor WT or Klf1(K74R) mice. Following BMT, bioluminescence imaging was performed on days 0, 10, 17 and 24.
Bone marrow transplantation
Request a detailed protocolBMT followed the standard protocol described in Imado et al., 2004. B6, CD45.1, or CD45.2 donor mice were sacrificed and their femurs were removed. Bone marrow cells were harvested by flashing the femurs with RPM I1640 medium (Gibco) using a 27-gauge needle and syringe. The cells were then incubated at 37°C for 30 min in murine complement buffer containing antibodies against B cells, T cells, and NK cells, washed twice with PBS, and then subjected to Ficoll-Paque PLUS gradient centrifugation to collect BMMNCs. BMMNCs (1 × 106 cells/mouse) from donor mice were injected into the tail veins of recipient B6, CD45.2, or CD45.1 mice that had been exposed to total body γ-irradiation of 10, 5, or 2.5 Gy.
Calcein-AM cytotoxicity assay of NK cells
Request a detailed protocol1 × 106 target cells (B16-F10 and Hepa1-6) were suspended in 1 ml DPBS containing 5 μM Calcein-AM (BioLegend) and incubated at 37°Cfor 30 min in the dark. The target cells were then washed by RPMI for three times and seeded at 0.3 × 105 cells per well on V-bottom 96-well plate. NK cells from the spleen were sorted by the surface markers, NK1.1+,CD3-, B220-, and NKp46+(Supplementary file 1a), centrifuged at 500 × g for 5 min, and cultured with IL-15 in RPMI. They were co-cultured with the target cells through different E (Effector):T (Target) ratios (0.5:1, 1:1, 3:1, and 9:1). The plates were centrifuged at 250 × g for 1 min to accelerate the contact of NK cells and target cells and then co-cultured for 4 hr at 37°C. Spontaneous release of Calcein-AM was measured in the target cells without NK cells, and the maximal release was determined by complete lysis of the target cells in RPMI containing 2% Triton-X 100. After co-culturing, the plates were centrifuged at 120 × g for 1 min and 100 μl/well of the supernatant was transferred to a 96-well assay plate (Corning). The 488/520 nm values were determined with EnSpire (PerkinElmer).
Bead-based multiplex assay of serum cytokines
Request a detailed protocolSerum samples were obtained via submandibular blood collection and allowed to clot in uncoated tubes for 2 hr at room temperature. The tubes were centrifuged at 6000 rpm and the supernatants were collected for cytokine analysis by bead-based multiplex assay (MILLIPLEX MAP Mouse High Sensitivity T Cell Panel, Millipore) following the manufacturer’s protocol (Aira et al., 2019).
Protein extraction
Request a detailed protocolThe cell pellets were resuspended in protein extraction buffer (20 mM HEPES, 0.2% SDS, 1 mM EDTA, 1 mM glycerophosphate, 1 mM Na3VO4, and 2.5 mM Na4P2O7) with protease inhibitor cocktail (Sigma-Aldrich) and 1 mM phenylmethylsulfonyl fluoride (PMSF). The lysates were further homogenized using a Bioruptor (Diagenode) at 4°C for 15 min, and then centrifuged at 14,000 × g at 4°C for 20 min. The supernatant was transferred to a new tube before determining protein concentration by means of BCA protein assay (Pierce, Thermo Fisher). Protein aliquots were stored at –30°C until use.
In-solution digestion
Request a detailed protocolProtein solutions were first diluted with 50 mM ammonium bicarbonate (ABC) and reduced with 5 mM dithiothreitol (DTT, Merck) at 60°C for 45 min, followed by cysteine-blocking with 10 mM iodoacetamide (IAM, Sigma) at 25°C for 30 min. The samples were then diluted with 25 mM ABC and digested with sequencing-grade modified porcine trypsin (Promega) at 37°C for 16 hr. The peptides were desalted using a homemade C18 microcolumn (SOURCE 15RPC, GE Healthcare) and stored at –30°C until use.
LC-MS/MS analysis
Request a detailed protocolThe desalted peptides were diluted in HPLC buffer A (0.1% formic acid in 30% acetonitrile) and loaded onto a homemade SCX column (0.6 × 5 mm, Luna 5 µm SCX 100 Å, Phenomenex). The eluted peptides were then trapped in a reverse-phase column (Zorbax 300 SB-C18, 0.3 × 5 mm; Agilent Technologies), and separated on a homemade column (HydroRP 2.5 µm, 75 μm I.D. × 15 cm with a 15 μm tip) using a multi-step gradient of HPLC buffer B (99.9% acetonitrile/0.1% formic acid) for 90 min with a flow rate of 0.3 μl/min. The LC apparatus was coupled to a 2D linear ion trap mass spectrometer (Orbitrap Elite ETD; Thermo Fisher) operated using Xcalibur 2.2 software (Thermo Fisher). Full-scan MS was performed in the Orbitrap over a range of 400–2000 Da and a resolution of 120,000 at m/z 400. Internal calibration was performed using the ion signal of [Si(CH3)2O]6H+ at m/z 536.165365 as lock mass. The 20 data-dependent MS/MS scan events were followed by one MS scan for the 20 most abundant precursor ions in the preview MS scan. The m/z values selected for MS/MS were dynamically excluded for 40 s with a relative mass window of 10 ppm. The electrospray voltage was set to 2.0 kV, and the temperature of the capillary was set to 200°C. MS and MS/MS automatic gain control was set to 1000 ms (full scan) and 200 ms (MS/MS), or to 3 × 106 ions (full scan) and 3,000 ions (MS/MS), for maximum accumulated time or ions, respectively.
Protein identification
Request a detailed protocolData analysis was carried out using Proteome Discoverer software (version 1.4, Thermo Fisher Scientific). The MS/MS spectra were searched against the SwissProt database using the Mascot search engine (Matrix Science, version 2.5). For peptide identification, 10 ppm mass tolerance was permitted for intact peptide masses, and 0.5 Da for CID fragment ions with an allowance for two missed cleavages arising from trypsin digestion, oxidized methionine and acetyl (protein N-terminal) as variable modifications, and carbamidomethyl (cysteine) as a static modification. Peptide spectrum matches (PSM) were then filtered based on high confidence and a Mascot search engine ranking of 1 for peptide identification to ensure an overall false discovery rate < 0.01. Proteins with single peptide hits were removed from further analysis.
Gene set enrichment analysis
Request a detailed protocolThe absolute abundance of each peptide was calculated from the respective peak intensity based on the PSM abundance. The protein abundance of each sample was calculated from the sum of the peptide abundance. The abundance data were then background-corrected and normalized according to variance stabilizing transformation by using the function ‘normalize_vsn’ in the R package DEP (Zhang et al., 2018). Differential expression across groups was determined using the function ‘test_diff’ based on protein-wise linear models combined with empirical Bayes statistics. Significant DEPs were determined according to a p-value threshold of 0.01 and a fold change > 1.5. To establish functional pathways enriched across groups, normalized data for each pair of compared groups were used to perform GSEA (version 4.2.0) (Subramanian et al., 2005) on selected MSigDB gene sets, including Hallmark (H), curated (C2), and immunologic signature (C7) gene sets, by using the default parameters. Normalized enrichment scores (NES) were used to plot a heatmap in the R package pheatmap (version 1.0.12).
Statistical analysis
Request a detailed protocolData are shown as mean ± SD or standard error of the mean (SEM). Comparisons of data under different experimental conditions were carried out using GraphPad Prism 6.0 software (GraphPad). Each error bar represents SEM unless otherwise indicated. Significant differences in tumor growth in mouse lungs were assessed using Student’s t-test. A difference between groups was considered statistically significant when the p-value was <0.05.
Data availability
All data that support the findings of this study are available from Dryad https://doi.org/10.5061/dryad.mkkwh717j.
-
Dryad Digital RepositoryLong-term Hematopoietic Transfer of the Anti-Cancer and Lifespan-Extending Capabilities of A Genetically Engineered Blood System by Transplantation of Bone Marrow Mononuclear Cells.https://doi.org/10.5061/dryad.mkkwh717j
References
-
Autophagy in healthy aging and diseaseNature Aging 1:634–650.https://doi.org/10.1038/s43587-021-00098-4
-
Engineering the next generation of cell-based therapeuticsNature Reviews. Drug Discovery 21:655–675.https://doi.org/10.1038/s41573-022-00476-6
-
Genes and longevity of lifespanInternational Journal of Molecular Sciences 23:1499.https://doi.org/10.3390/ijms23031499
-
Biomimetic quantum dot-labeled B16F10 murine melanoma cells as a tool to monitor early steps of lung metastasis by in vivo imagingInternational Journal of Nanomedicine 13:6391–6412.https://doi.org/10.2147/IJN.S165565
-
Bone marrow chimeras-a vital tool in basic and translational researchJournal of Molecular Medicine 97:889–896.https://doi.org/10.1007/s00109-019-01783-z
-
Umbilical cord blood and serum for the treatment of ocular diseases: A comprehensive reviewOphthalmology and Therapy 9:235–248.https://doi.org/10.1007/s40123-020-00239-9
-
Immune modulatory effects of oncogenic KRAS in cancerNature Communications 11:5439.https://doi.org/10.1038/s41467-020-19288-6
-
Clinical serum therapy: Benefits, cautions, and potential applicationsThe Keio Journal of Medicine 66:57–64.https://doi.org/10.2302/kjm.2016-0017-IR
-
Negative regulation of the differentiation of flk2− cd34− lsk hematopoietic stem cells by EKLF/KLF1International Journal of Molecular Sciences 21:8448.https://doi.org/10.3390/ijms21228448
-
Cancer immunotherapies targeting the PD-1 signaling pathwayJournal of biomedical science 24:26.https://doi.org/10.1186/s12929-017-0329-9
-
Activation and repression of interleukin-12 p40 transcription by erythroid Kruppel-like factor in macrophagesThe Journal of Biological Chemistry 279:18451–18456.https://doi.org/10.1074/jbc.M400320200
-
Cellular and molecular mechanisms involved in hematopoietic stem cell aging as a clinical prospectOxidative Medicine and Cellular Longevity 2022:2713483.https://doi.org/10.1155/2022/2713483
-
Interferon gamma in cancer immunotherapyCancer Medicine 7:4509–4516.https://doi.org/10.1002/cam4.1700
-
TGF-β: duality of function between tumor prevention and carcinogenesisJournal of the National Cancer Institute 106:djt369.https://doi.org/10.1093/jnci/djt369
-
Age and age-related diseases: Role of inflammation triggers and cytokinesFrontiers in Immunology 9:586.https://doi.org/10.3389/fimmu.2018.00586
-
Negative regulation of natural killer cell function by EAT-2, a SAP-related adaptorNature Immunology 6:1002–1010.https://doi.org/10.1038/ni1242
-
Hematopoietic stem cell: self-renewal versus differentiationWiley Interdisciplinary Reviews. Systems Biology and Medicine 2:640–653.https://doi.org/10.1002/wsbm.86
-
Sumoylation of EKLF promotes transcriptional repression and is involved in inhibition of megakaryopoiesisMolecular and Cellular Biology 27:8547–8560.https://doi.org/10.1128/MCB.00589-07
-
Novel roles for KLF1 in erythropoiesis revealed by mRNA-seqGenome research 22:2385–2398.https://doi.org/10.1101/gr.135707.111
-
Interleukin-4 receptor signaling and its binding mechanism: A therapeutic insight from inhibitors tool boxCytokine & growth factor reviews 32:3–15.https://doi.org/10.1016/j.cytogfr.2016.04.002
-
Clinical application of intermittent fasting for weight loss: progress and future directionsNature Reviews. Endocrinology 18:309–321.https://doi.org/10.1038/s41574-022-00638-x
-
Morphine promotes migration and lung metastasis of mouse melanoma cellsBrazilian Journal of Anesthesiology 73:441–445.https://doi.org/10.1016/j.bjane.2021.10.019
-
Natural killer cells in cancer biology and therapyMolecular Cancer 19:120.https://doi.org/10.1186/s12943-020-01238-x
-
Hypoxia and agingExperimental & molecular medicine 51:1–15.https://doi.org/10.1038/s12276-019-0233-3
-
Proteome-wide identification of ubiquitin interactions using UbIA-MSNature Protocols 13:530–550.https://doi.org/10.1038/nprot.2017.147
-
Proteomic profiling of lymphocytes in autoimmunity, inflammation and cancerJournal of Translational Medicine 12:6.https://doi.org/10.1186/1479-5876-12-6
-
Inflammation, epigenetics, and metabolism converge to cell senescence and ageing: the regulation and interventionSignal Transduction and Targeted Therapy 6:245.https://doi.org/10.1038/s41392-021-00646-9
-
Type I interferons in anticancer immunityNature Reviews. Immunology 15:405–414.https://doi.org/10.1038/nri3845
Article and author information
Author details
Funding
Taipei Medical University
- C-K James Shen
Academia Sinica
- C-K James Shen
Chang Gung Memorial Hospital (CMRPG2L0081)
- Yu-Chiau Shyu
Chang Gung Memorial Hospital (CMRPG2G0311)
- Yu-Chiau Shyu
National Science and Technology Council (NSC 102-2320-B-001-010)
- Nan-Shih Liao
National Science and Technology Council (MOST 103-2311-B-010-003)
- Yu-Chiau Shyu
National Science and Technology Council (MOST 110-2628-B-001-024)
- Liuh-Yow Chen
The funders had no role in study design, data collection and interpretation, or the decision to submit the work for publication.
Acknowledgements
We thank Ya-Min Lin and her colleagues at the FACS Core for their efforts in cell sorting. We also thank Taiwan Mouse Clinic facility for their expert help in the IVIS experiments. The effort by Dr. Ching-Yen Tsai at the Transgenic Core Facility, IMB, is greatly appreciated. The Immune Monitoring Core of Taipei Medical University provided great help in the analysis of the blood cell populations. We also thank the Bioinformatics Core Facility, IMB, for its efforts on the proteomics analysis, and Dr. Da-Liang Ou at the National Taiwan University for providing us with the Hepa1-6 cell line. This work was supported by Taipei Medical University (to CKJS), Academia Sinica (to CKJS), Chang-Gung Memorial Hospital Research Project (CMRPG2L0081 and CMRPG2G0311to YCS), and grants from the National Science and Technology Council, Taiwan, ROC (NSC 102-2320-B-001-010 to NSL, MOST 103-2311-B-010-003 YCS, and MOST 110-2628-B-001-024 to LYC).
Ethics
All the animal procedures were approved by the Institute of Animal Care and Use Committees, IACUC, at Academia Sinica. The IACUC numbers are 17-02-1052, 17-12-1142, 20-10-1528, BioTReC-110-M-005, and BioTReC-111-M-004, respectively.
Version history
Cite all versions
You can cite all versions using the DOI https://doi.org/10.7554/eLife.88275. This DOI represents all versions, and will always resolve to the latest one.
Copyright
© 2023, Wang, Hung et al.
This article is distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use and redistribution provided that the original author and source are credited.
Metrics
-
- 825
- views
-
- 83
- downloads
-
- 0
- citations
Views, downloads and citations are aggregated across all versions of this paper published by eLife.
Download links
A two-part list of links to download the article, or parts of the article, in various formats.
Downloads (link to download the article as PDF)
Open citations (links to open the citations from this article in various online reference manager services)
Cite this article (links to download the citations from this article in formats compatible with various reference manager tools)
Long-term hematopoietic transfer of the anti-cancer and lifespan-extending capabilities of a genetically engineered blood system by transplantation of bone marrow mononuclear cells
eLife 12:RP88275.
https://doi.org/10.7554/eLife.88275.3
Further reading
-
- Stem Cells and Regenerative Medicine
Tissue engineering strategies predominantly rely on the production of living substitutes, whereby implanted cells actively participate in the regenerative process. Beyond cost and delayed graft availability, the patient-specific performance of engineered tissues poses serious concerns on their clinical translation ability. A more exciting paradigm consists in exploiting cell-laid, engineered extracellular matrices (eECMs), which can be used as off-the-shelf materials. Here, the regenerative capacity solely relies on the preservation of the eECM structure and embedded signals to instruct an endogenous repair. We recently described the possibility to exploit custom human stem cell lines for eECM manufacturing. In addition to the conferred standardization, the availability of such cell lines opened avenues for the design of tailored eECMs by applying dedicated genetic tools. In this study, we demonstrated the exploitation of CRISPR/Cas9 as a high precision system for editing the composition and function of eECMs. Human mesenchymal stromal/stem cell (hMSC) lines were modified to knock out vascular endothelial growth factor (VEGF) and Runt-related transcription factor 2 (RUNX2) and assessed for their capacity to generate osteoinductive cartilage matrices. We report the successful editing of hMSCs, subsequently leading to targeted VEGF and RUNX2-knockout cartilage eECMs. Despite the absence of VEGF, eECMs retained full capacity to instruct ectopic endochondral ossification. Conversely, RUNX2-edited eECMs exhibited impaired hypertrophy, reduced ectopic ossification, and superior cartilage repair in a rat osteochondral defect. In summary, our approach can be harnessed to identify the necessary eECM factors driving endogenous repair. Our work paves the road toward the compositional eECMs editing and their exploitation in broad regenerative contexts.
-
- Developmental Biology
- Stem Cells and Regenerative Medicine
In holothurians, the regenerative process following evisceration involves the development of a ‘rudiment’ or ‘anlage’ at the injured end of the mesentery. This regenerating anlage plays a pivotal role in the formation of a new intestine. Despite its significance, our understanding of the molecular characteristics inherent to the constituent cells of this structure has remained limited. To address this gap, we employed state-of-the-art scRNA-seq and hybridization chain reaction fluorescent in situ hybridization analyses to discern the distinct cellular populations associated with the regeneration anlage. Through this approach, we successfully identified 13 distinct cell clusters. Among these, two clusters exhibit characteristics consistent with putative mesenchymal cells, while another four show features akin to coelomocyte cell populations. The remaining seven cell clusters collectively form a large group encompassing the coelomic epithelium of the regenerating anlage and mesentery. Within this large group of clusters, we recognized previously documented cell populations such as muscle precursors, neuroepithelial cells, and actively proliferating cells. Strikingly, our analysis provides data for identifying at least four other cellular populations that we define as the precursor cells of the growing anlage. Consequently, our findings strengthen the hypothesis that the coelomic epithelium of the anlage is a pluripotent tissue that gives rise to diverse cell types of the regenerating intestinal organ. Moreover, our results provide the initial view into the transcriptomic analysis of cell populations responsible for the amazing regenerative capabilities of echinoderms.




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}