Synthetic analysis of trophic diversity and evolution in Enantiornithes with new insights from Bohaiornithidae
- Department of Earth Sciences, The University of Hong Kong, China
- School of Natural Sciences, University of Hull, United Kingdom
- Institute of Geology and Paleontology, Linyi University, China
- Shandong Tianyu Museum of Nature, China
- School of Life Sciences, The Chinese University of Hong Kong, China
Figures
Figure 1

Life reconstruction of enantiornithine birds feeding.
Longipteryx (left), Bohaiornis (centre), and Pengornis (right) are pictured in the Early Cretaceous forests of northeastern China, roughly 120 million years ago. Bohaiornis is depicted feeding on cypress (Cupressaceae, Ding et al., 2016) leaves after the findings in this work. Longipteryx is depicted feeding on the mayfly Epicharmeropsis hexavenulosus (Huang et al., 2007) after (Miller et al., 2022). Pengornis is depicted feeding on the fish Lycoptera davidi (Chang and Miao, 2004) after Miller et al., 2023.
Figure 2 with 1 supplement
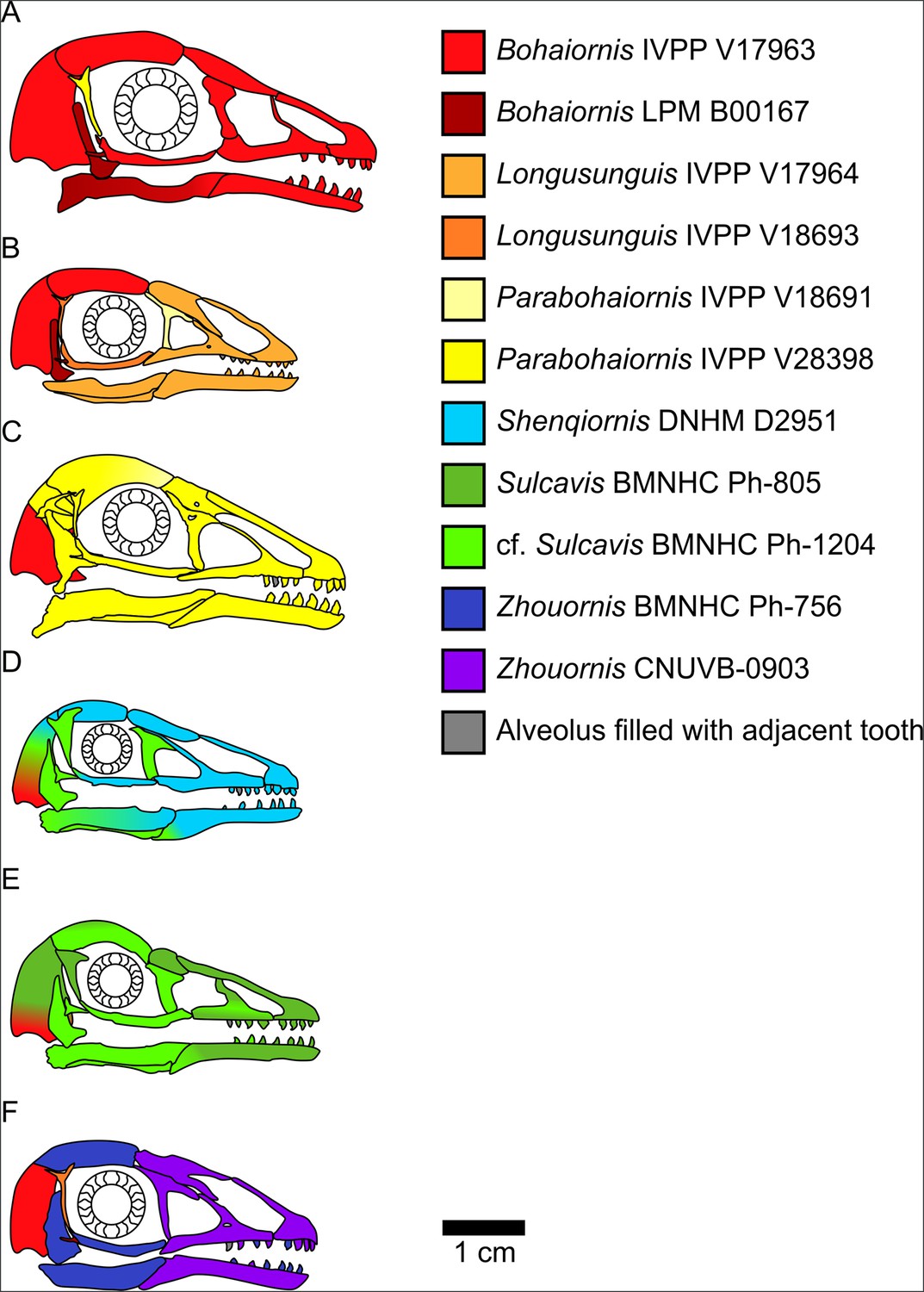
Bohaiornithid skull reconstructions used for mechanical advantage (MA) and finite element analysis (FEA) calculations in this study.
Reconstructions are of Bohaiornis (A), Longusunguis (B), Parabohaiornis (C), Shenqiornis (D), Sulcavis (E), and Zhouornis (F). Different colours indicate elements taken from different individual specimens. All sclerotic rings are based on Longipteryx specimen BMNHC Ph-930B. See the ‘Methods’ section for more details on reconstruction. Scale for each reconstruction is based on the individual which makes up the largest portion of the reconstruction.
Figure 2—figure supplement 1
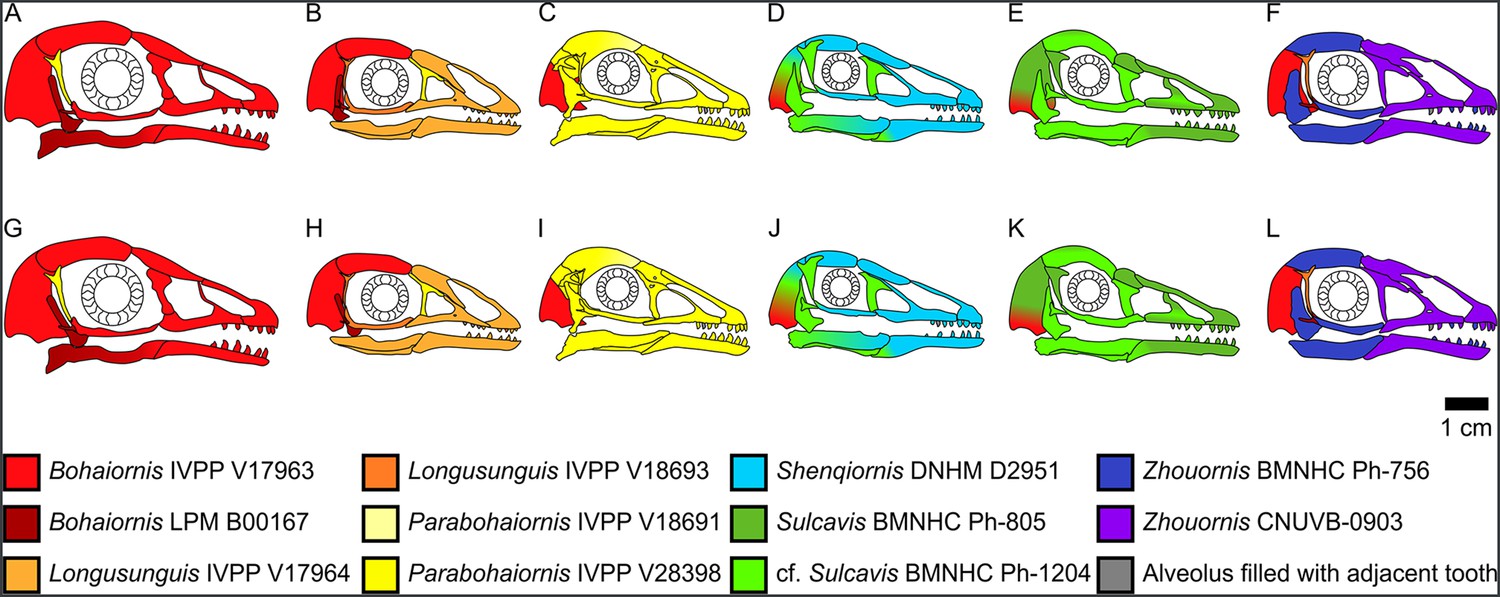
Reconstructions of bohaiornithid skulls for sensitivity analysis of the quadrate’s position, which is uncertain due to disarticulation in all specimens.
Reconstructions of Bohaiornis (A, G), Longusunguis (B, H), Parabohaiornis (C, I), Shenqiornis (D, J), Sulcavis (E, K), and Zhouornis (F, L) are constructed with the quadrate shifted as far anteriorly (A–F) or posteriorly (G–L) as biologically plausible.
Figure 3
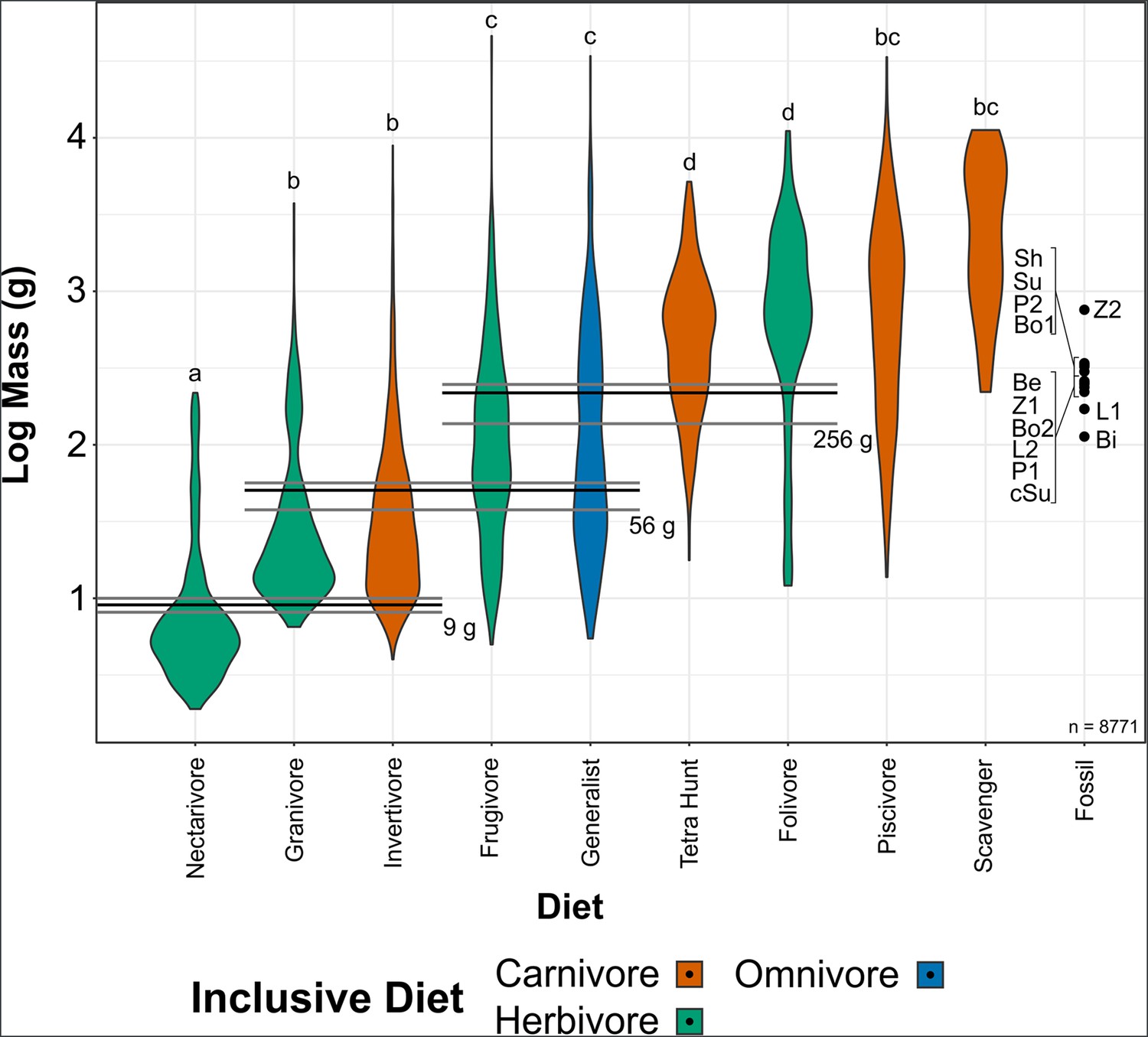
Violin plots of bird mass by diet, arranged in order of ascending mean mass.
Masses were tested for significant differences via phylogenetic honest significant differences (HSD). Diets marked with the same letter are not significantly different from one another. Cut-off points between significantly different mass groups (black lines, with 95% CIs as grey lines) were calculated by optimising the Youden index and plotted. Note that, unlike in other diet treatments herein, granivores are not separated into husking and swallowing granivores. Mean bohaiornithid mass estimates are plotted for comparison, see Table 3. Diet abbreviations: Tetra Hunt, Tetrapod Hunter. Fossil taxon abbreviations: Be, Beiguornis khinganensis MHGU-F307/8; Bi, Bohaiornithidae indet. CUGB P1202; Bo1, Bohaiornis LPM B00167; Bo2, Bohaiornis IVPP V17963; L1, Longusunguis IVPP V17964; L2, Longusunguis IVPP V18693; P1, Parabohaiornis IVPP V18691; P2, Parabohaiornis IVPP V28398; Sh, Shenqiornis DNHM D2950/1; Su, Sulcavis BMNH Ph-805; cSu, cf. Sulcavis BMNHC-Ph1204; Z1, Zhouornis CNUVB-0903, Z2, Zhouornis BMNHC Ph 756.
Figure 4 with 2 supplements
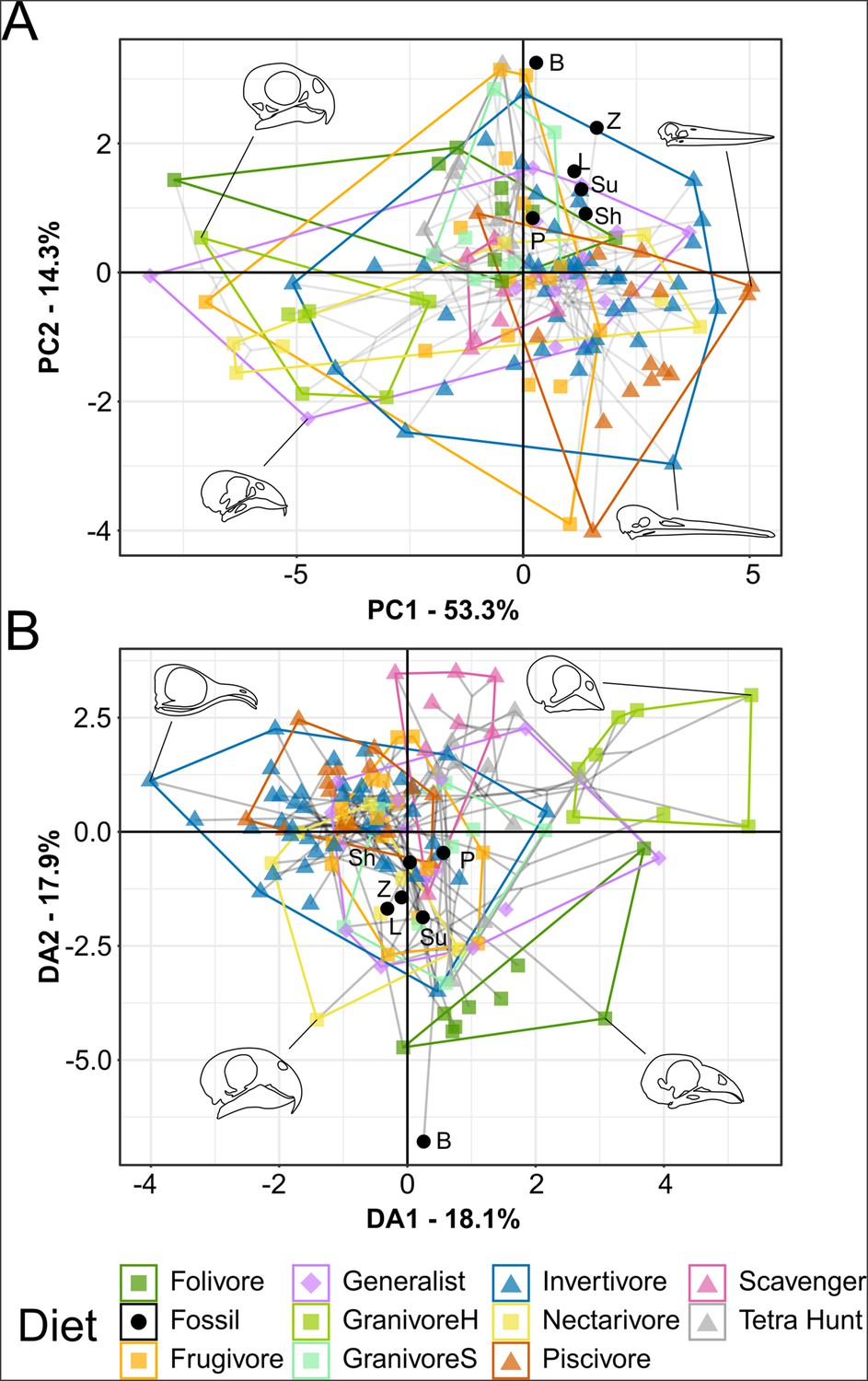
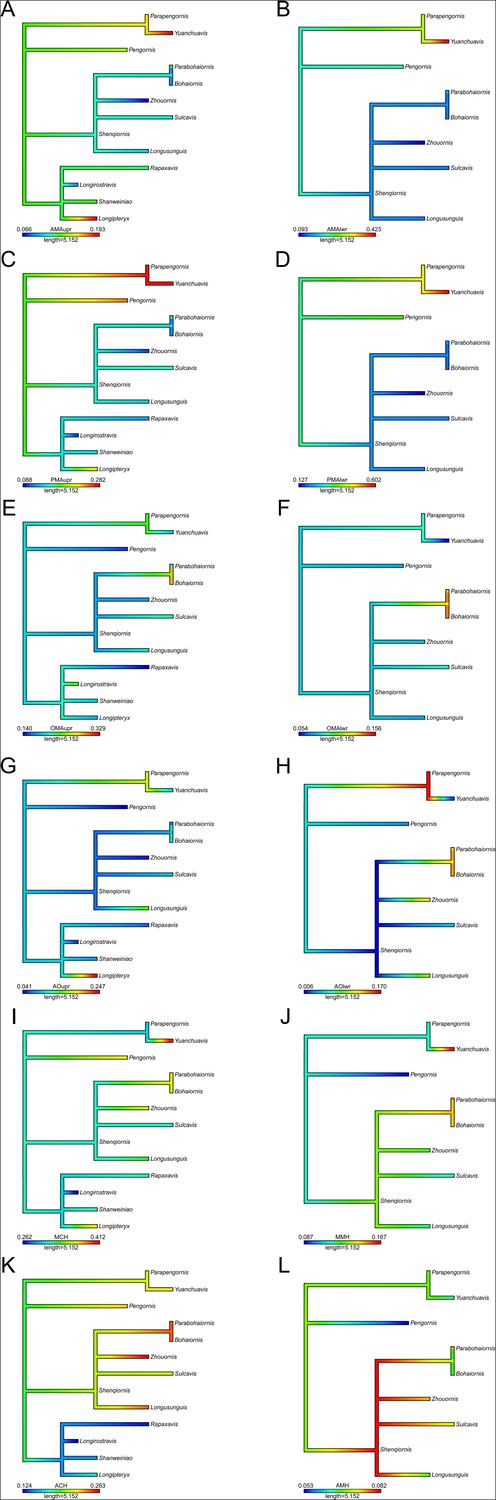
Functional phylomorphospace of mechanical advantage (MA) and functional index data, grouped by diet.
Grey lines indicate phylogenetic relationships. Line drawings of skulls for selected taxa are provided for reference. Data are presented with principal component analysis (PCA) (A) and flexible discriminant analysis (FDA) (B). See Figure 4—figure supplement 1 for character weights and Table 4 for FDA posterior predictions. Diet abbreviations: GranivoreH, Husking Granivore; GranivoreS, Swallowing Granivore; Tetra Hunt, Tetrapod Hunter. Fossil taxon abbreviations: B, Bohaiornis; L, Longusunguis; P, Parabohaiornis; Sh, Shenqiornis; Su, Sulcavis; Z, Zhouornis.
Figure 4—figure supplement 1
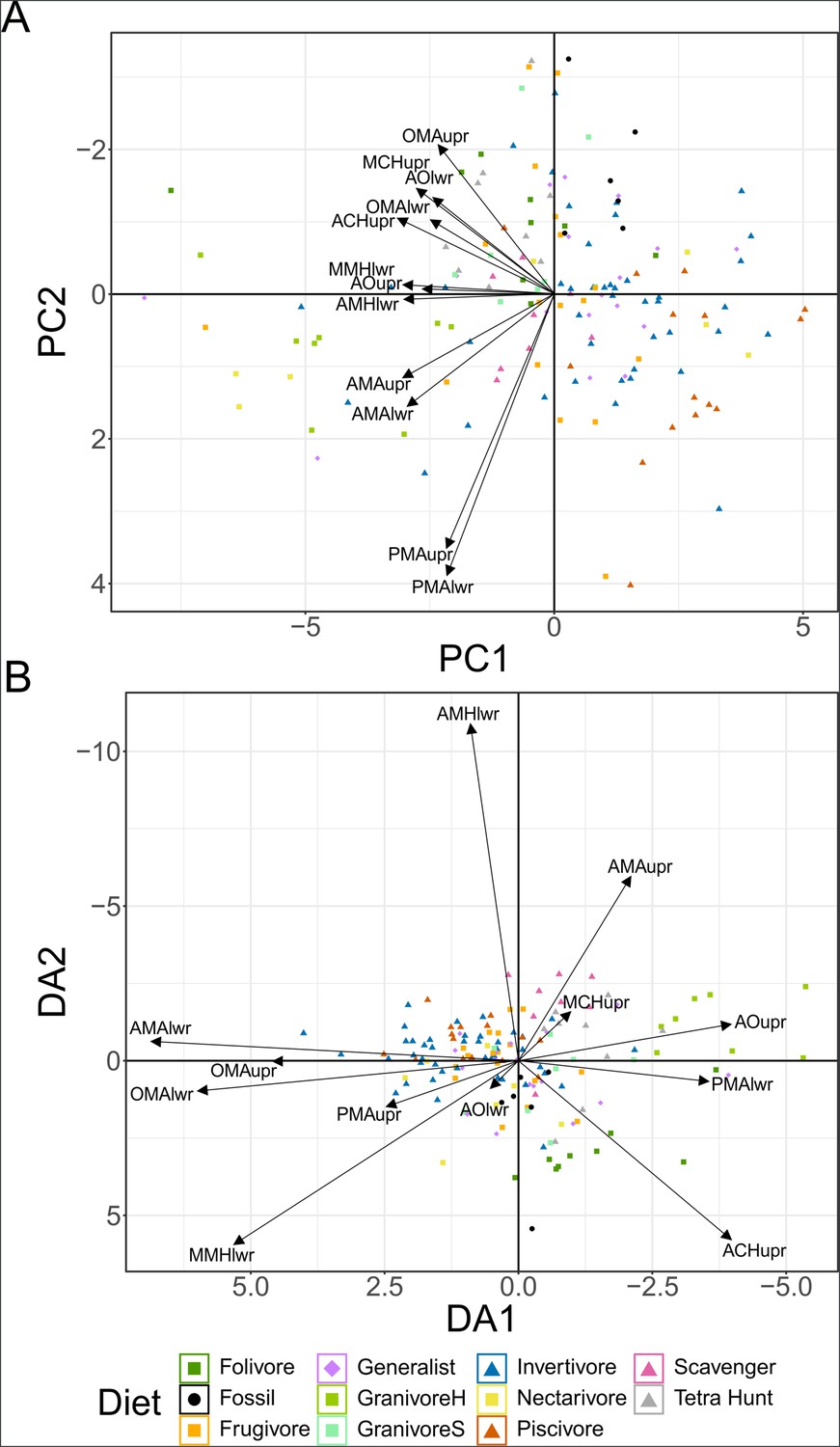
Plot of character weightings for the graphs in Figure 4.
Plots are provided for principal component analysis (PCA) (A) and flexible discriminate analysis (FDA) (B). Character abbreviations: AMA, anterior jaw-closing mechanical advantage; PMA, posterior jaw-closing mechanical advantage; OMA, jaw-opening mechanical advantage; AO, relative articular offset; MCH, relative maximum cranial height; MMH, relative maximum mandible height; ACH, relative average cranial height; AMH, relative average mandible height. Suffixes of upr and lwr respectively denote the measurement is taken of the upper or lower jaw. Diet abbreviations: GranivoreH, Husking Granivore; GranivoreS, Swallowing Granivore; Tetra Hunt, Tetrapod Hunter.
Figure 4—figure supplement 2
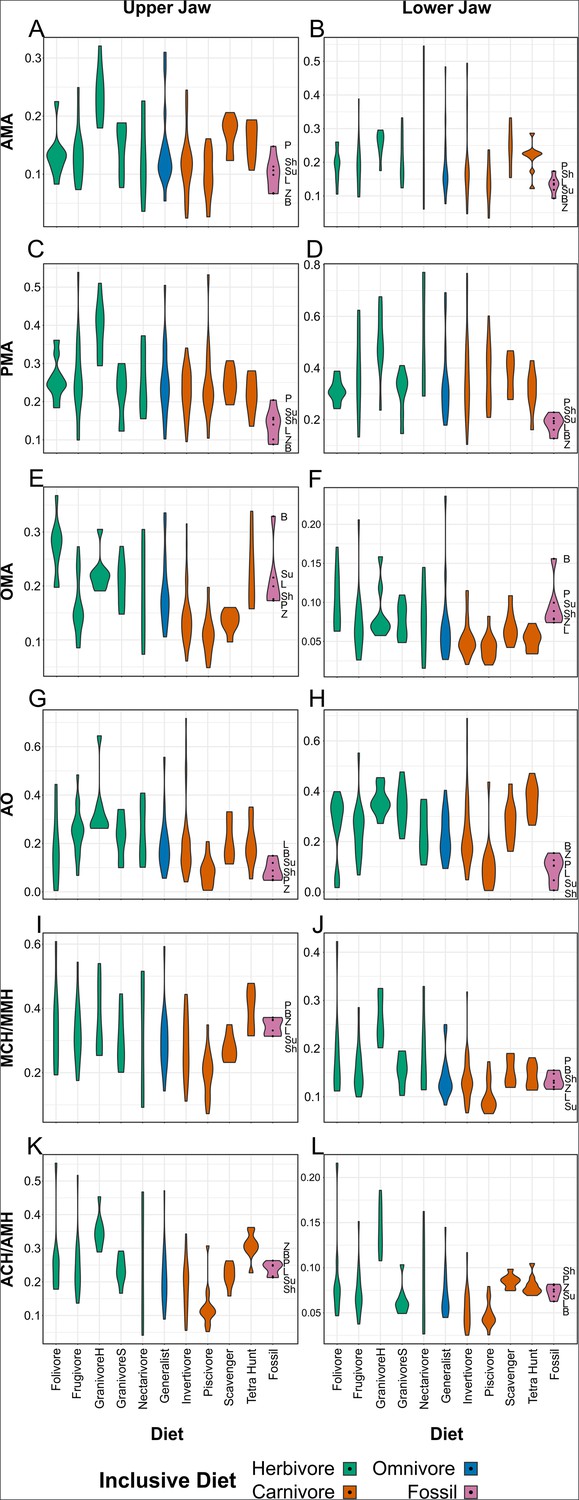
Violin plots of individual functional indices, organised by diet.
AMA (A, B), PMA (C, D), OMA (E, F), AO (G, H), MCH (I), MMH (J), ACH (K), and AMH (L) values are provided for the upper (A, C, E, G, I, K) and lower (B, D, F, H, J, L) jaws. Character abbreviations: AMA, anterior jaw-closing mechanical advantage; PMA, posterior jaw-closing mechanical advantage; OMA, jaw-opening mechanical advantage; AO, relative articular offset; MCH, relative maximum cranial height; MMH, relative maximum mandible height; ACH, relative average cranial height; AMH, relative average mandible height. Diet abbreviations: GranivoreH, Husking Granivore; GranivoreS, Swallowing Granivore; Tetra Hunt, Tetrapod Hunter. Fossil taxon abbreviations: B, Bohaiornis; L, Longusunguis; P, Parabohaiornis; Sh, Shenqiornis; Su, Sulcavis; Z, Zhouornis.
Figure 5
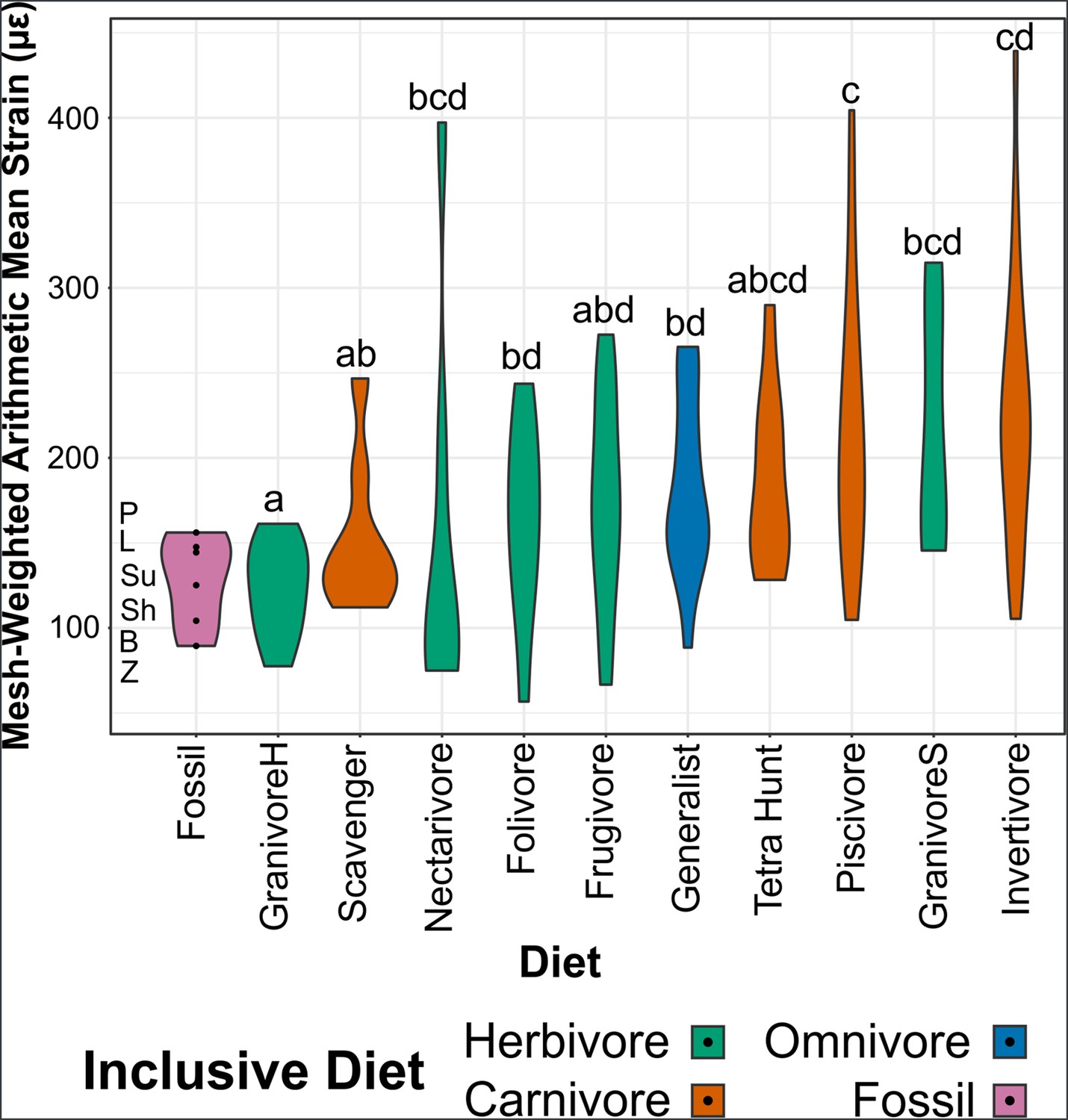
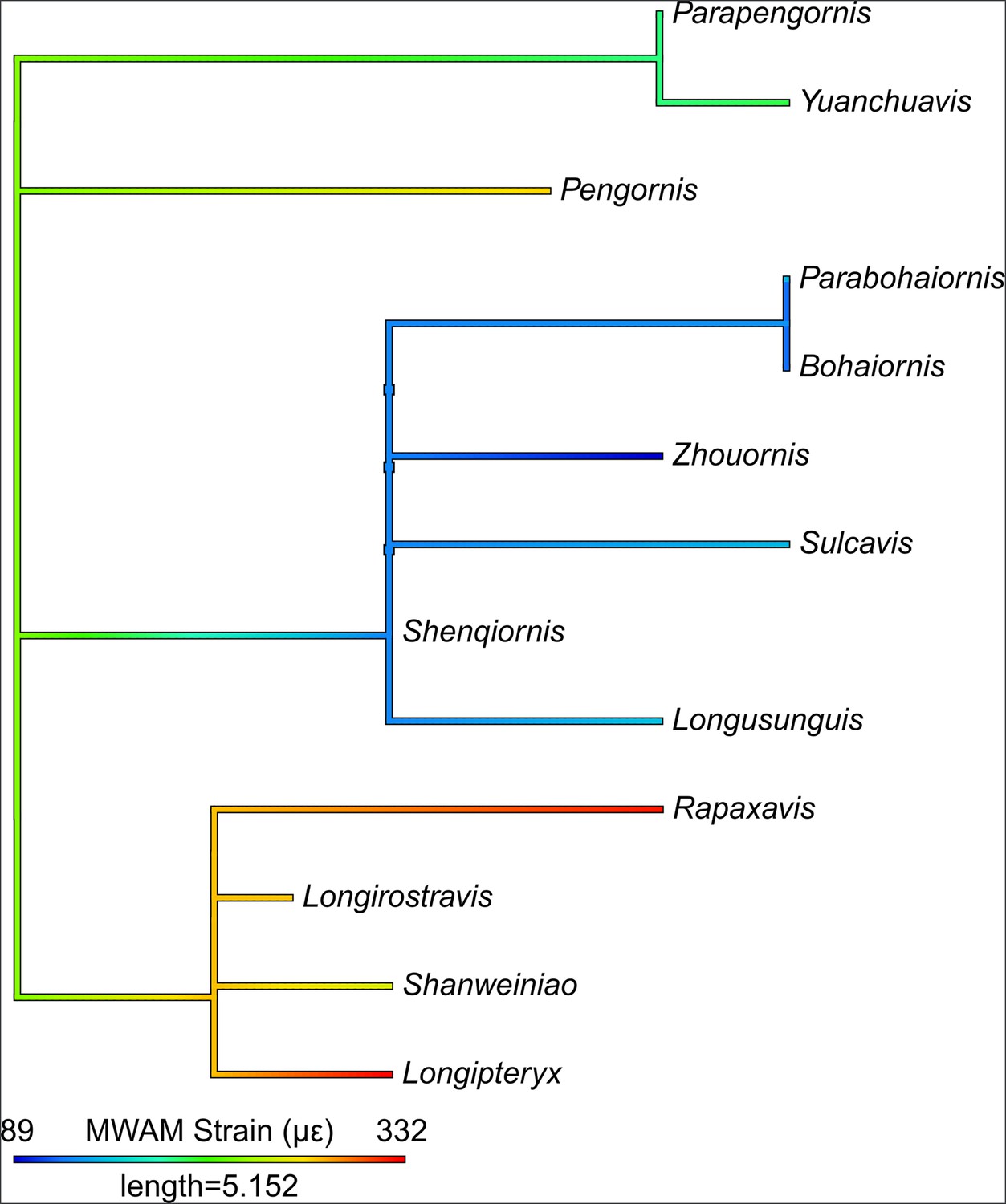
Violin plots of mesh-weighted arithmetic mean (MWAM) strain of finite element analysis (FEA) models, organised by diet.
Extant diets ascend in average MWAM strain from left to right. MWAM strains were tested for significant differences via phylogenetic honest significant differences (HSD). Diets marked with the same letter are not significantly different from one another. Diet abbreviations: GranivoreH, Husking Granivore; GranivoreS, Swallowing Granivore; Tetra Hunt, Tetrapod Hunter. Fossil taxon abbreviations: B, Bohaiornis; L, Longusunguis; P, Parabohaiornis; Sh, Shenqiornis; Su, Sulcavis; Z, Zhouornis.
Figure 6
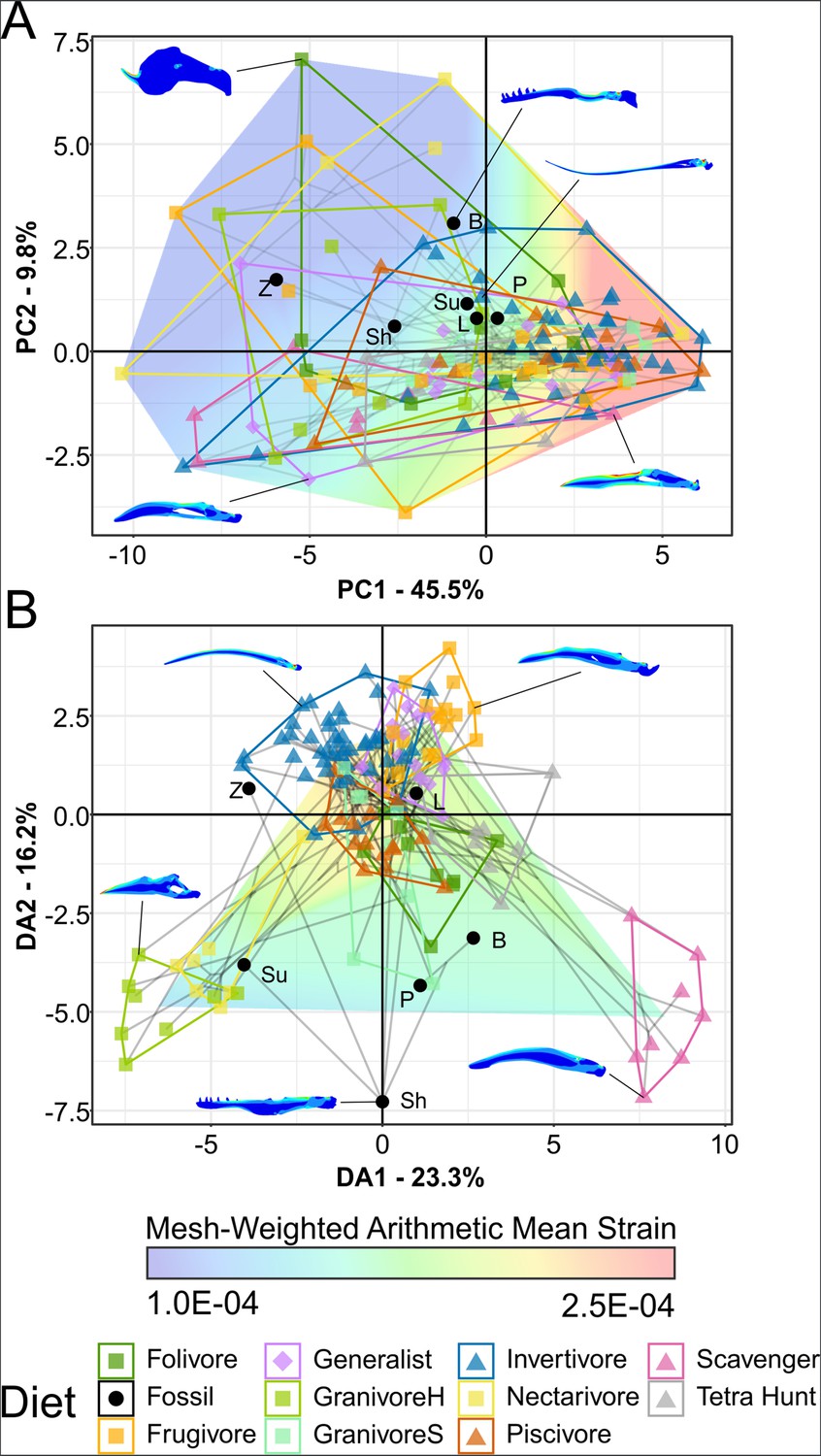
Phylogenetic strain-space of total maximum in-plane principal strain intervals for extant and fossil bird lower jaw finite element models, grouped by diet.
Mesh-weighted arithmetic mean (MWAM) strain is mapped overtop the data. Grey lines indicate phylogenetic relationships. Contour plots for selected taxa are provided for reference. Data are presented with principal component analysis (PCA) (A) and flexible discriminate analysis (FDA) (B). In PCA (A), overall strain increases along PC1, and strain heterogeneity (i.e. lower areas of intermediate strain) increases along PC2. In FDA (B), DA1 and DA2 have loadings of various similar low-strain intervals, with high-strain intervals clustering near the origin. See Table 5 for FDA posterior predictions. Diet abbreviations: GranivoreH, Husking Granivore; GranivoreS, Swallowing Granivore; Tetra Hunt, Tetrapod Hunter. Fossil taxon abbreviations: B, Bohaiornis; L, Longusunguis; P, Parabohaiornis; Sh, Shenqiornis; Su, Sulcavis; Z, Zhouornis.
Figure 7 with 2 supplements
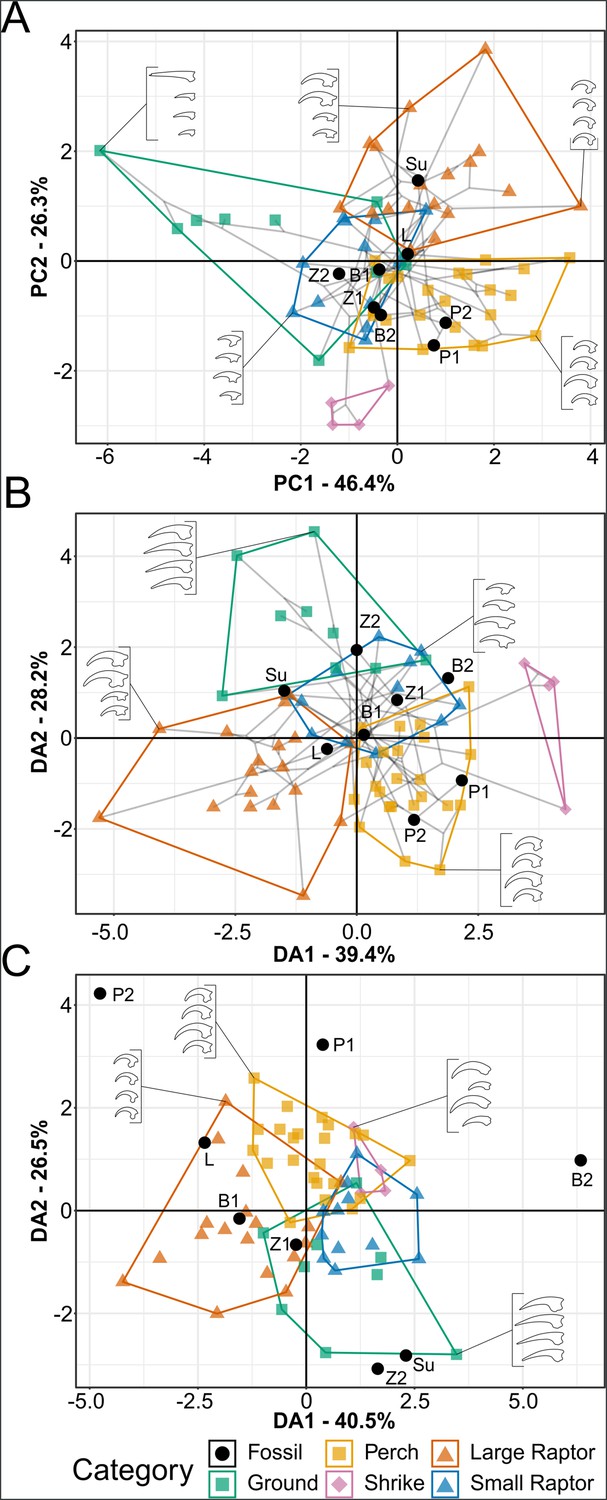
Phylomorphospace of extant and fossil bird claw shape from pedal traditional morphometric (TM), grouped by pedal ecological category.
Grey lines indicate phylogenetic relationships. Line drawings of claws for selected taxa are provided for reference. Data are presented with principal component analysis (PCA) (A), flexible discriminate analysis (FDA) (B), and phylogenetic flexible discriminate analysis (pFDA) (C). See Figure 7—figure supplement 1 for character weights and Table 6 for FDA and pFDA posterior predictions. Category abbreviations: large raptor, raptor taking prey which does not fit in the foot; small raptor, raptor taking prey which can fit in the foot. Fossil taxon abbreviations: B1, Bohaiornis LPM B00167; B2, Bohaiornis IVPP V17963; L, Longusunguis IVPP V18693; P1, Parabohaiornis IVPP V18690; P2, Parabohaiornis IVPP V18691; Su, Sulcavis BMNH Ph-805; Z1, Zhouornis CNUVB-0903, Z2, Zhouornis BMNHC Ph 756.
Figure 7—figure supplement 1
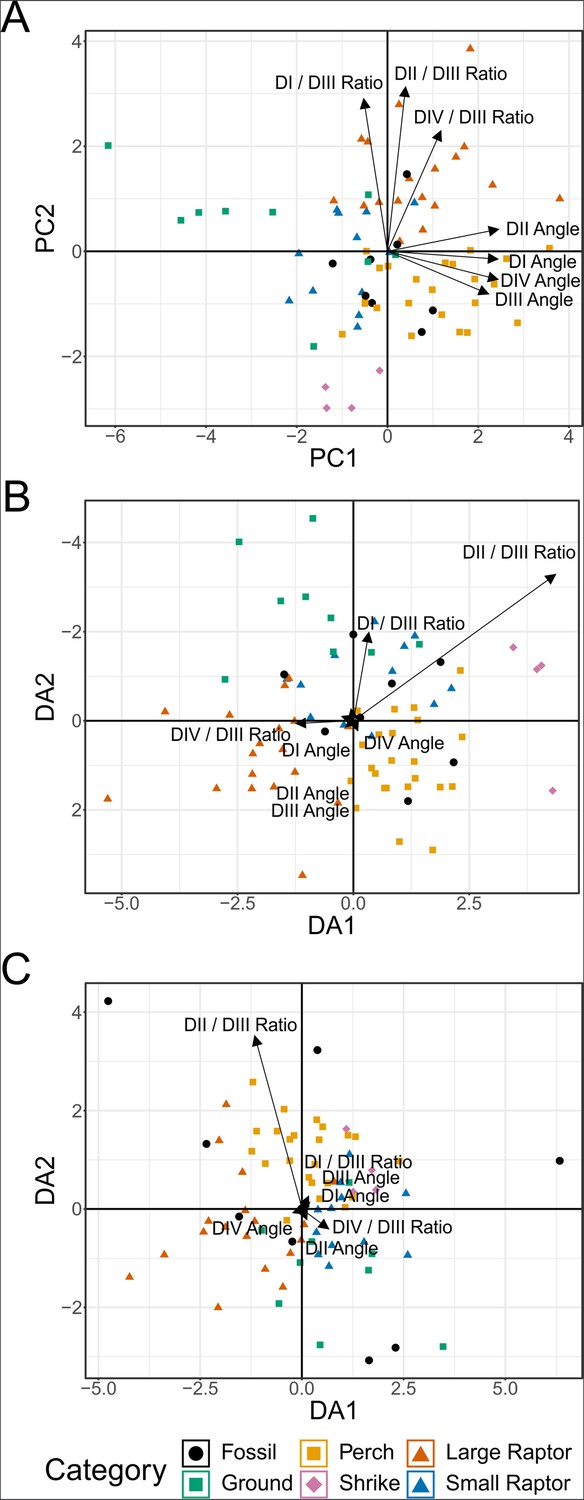
Plot of character weightings for the graphs in Figure 7.
Plots are provided for principal component analysis (PCA) (A), flexible discriminate analysis (FDA) (B), and phylogenetic flexible discriminate analysis (pFDA) (C). Category abbreviations: large raptor, raptor taking prey which does not fit in the foot; small raptor, raptor taking prey which can fit in the foot.
Figure 7—figure supplement 2
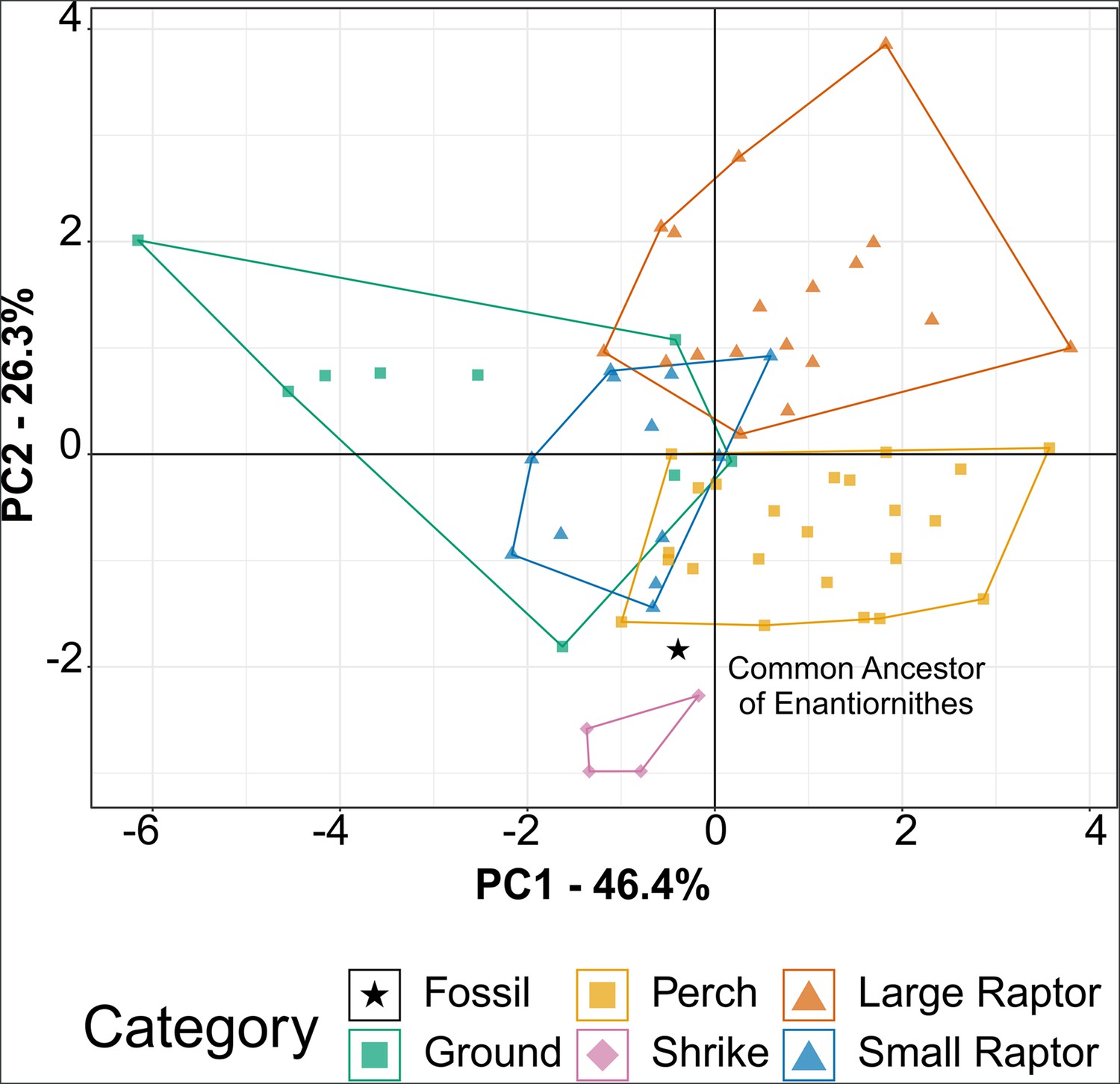
Phylomorphospace of extant bird claw shape from traditional morphometric (TM), grouped by pedal ecological category, for comparison to the common ancestor of Enantiornithes.
See Figure 7—figure supplement 1A for character weights. Category abbreviations: large raptor, raptor taking prey which does not fit in the foot; small raptor, raptor taking prey which can fit in the foot.
Figure 8 with 1 supplement
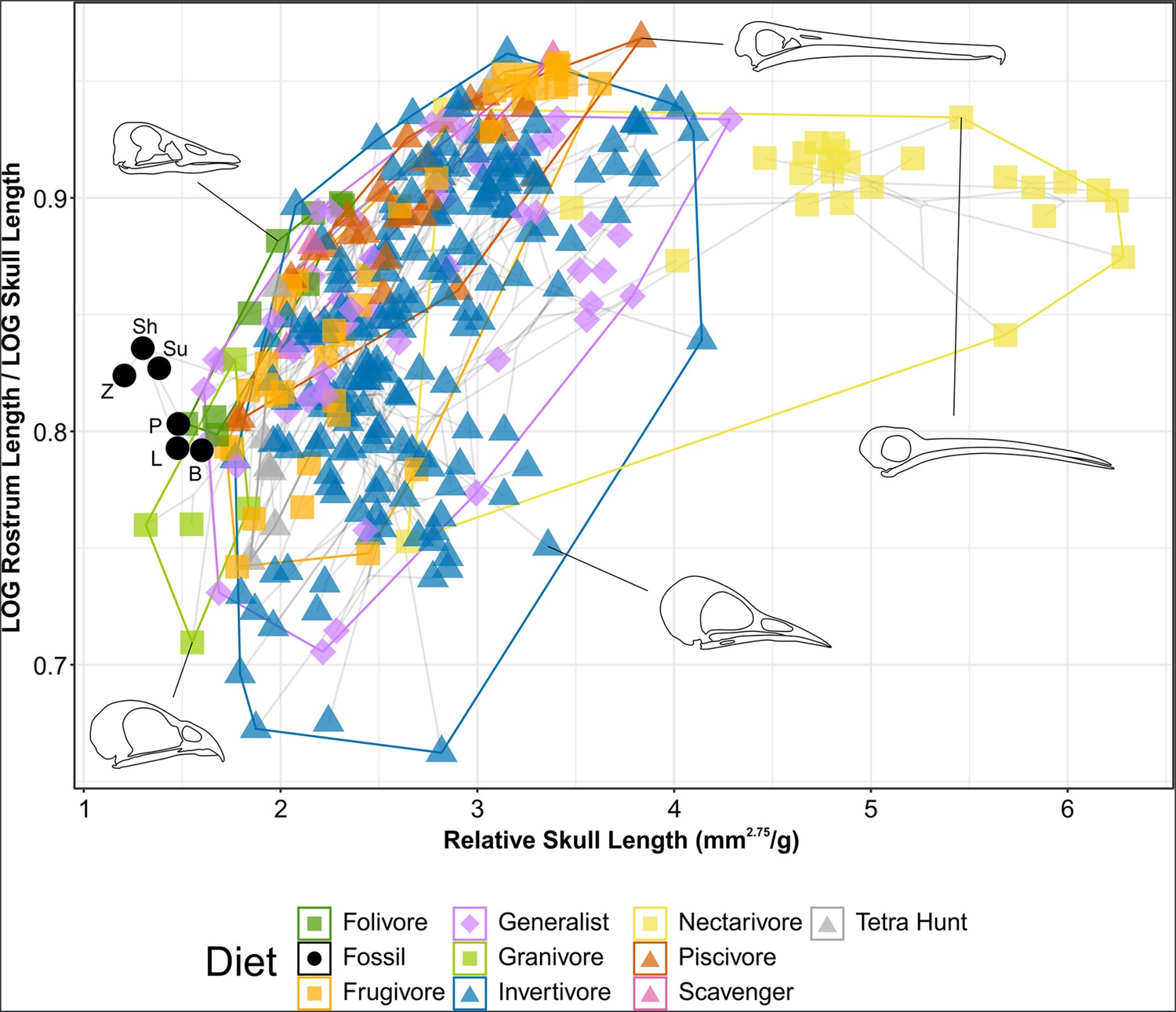
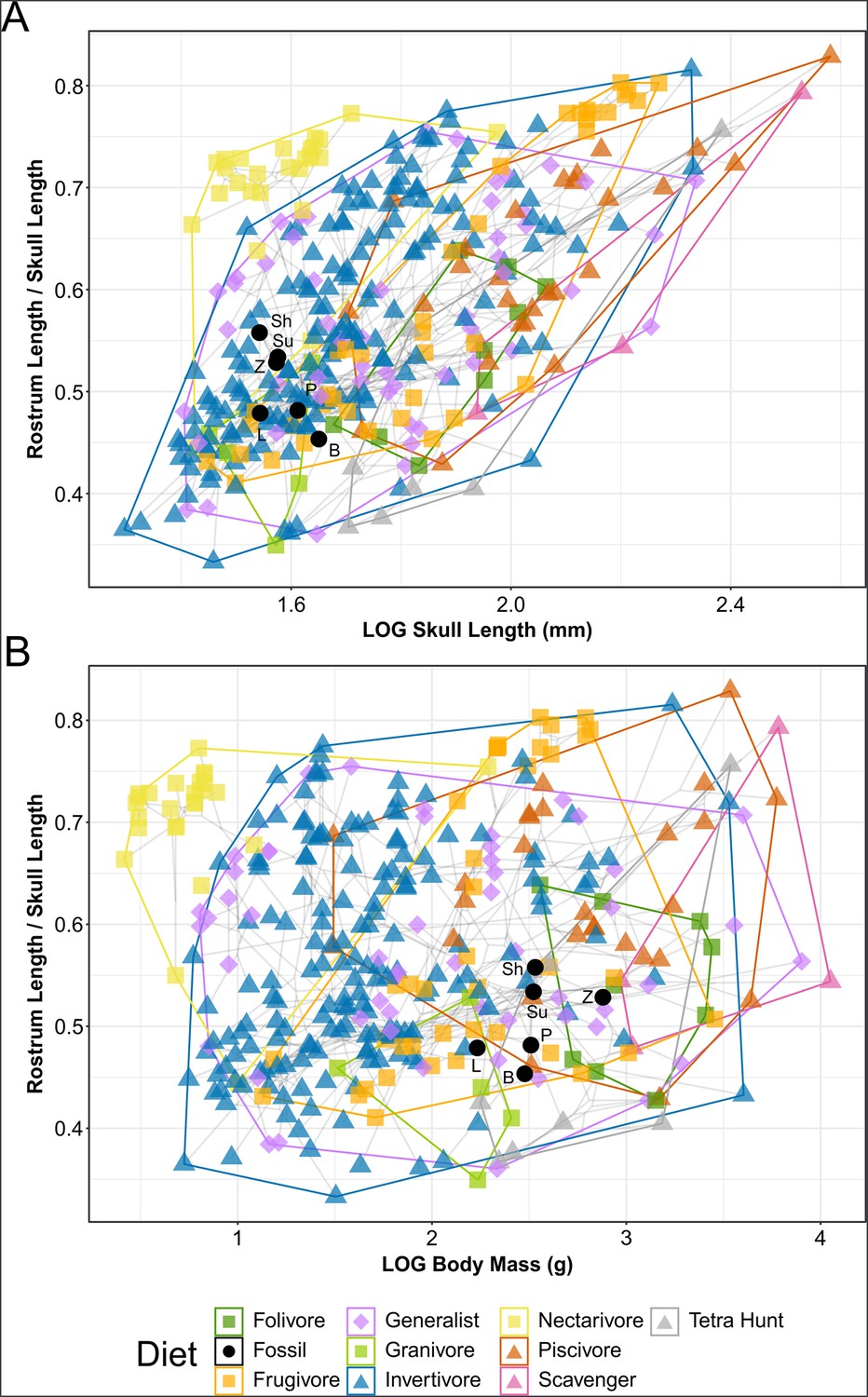
Phylomorphospace of extant and fossil bird skull proportions, grouped by diet.
Grey lines indicate phylogenetic relationships. Line drawings of skulls for selected taxa are provided for reference. The data presented are modified from Clark et al., 2023, see Figure 8—figure supplement 1 for data more directly comparable to that study. Diet abbreviation: Tetra Hunt, Tetrapod Hunter. Fossil taxon abbreviations: B, Bohaiornis; L, Longusunguis; P, Parabohaiornis; Sh, Shenqiornis; Su, Sulcavis; Z, Zhouornis.
Figure 8—figure supplement 1
Alternate phylomorphospaces to Figure 8.
Grey lines indicate phylogenetic relationships. Axes are presented as shown in Clark et al., 2023 (A) and with mass as the x-axis to investigate if spreading is driven by size effects (B). As we interpret these trends as being size-dependent, most discussion of skull proportions in this work are based on the main figure. Diet abbreviation: Tetra Hunt, Tetrapod Hunter. Fossil taxon abbreviations: B, Bohaiornis; L, Longusunguis; P, Parabohaiornis; Sh, Shenqiornis; Su, Sulcavis; Z, Zhouornis.
Figure 9 with 4 supplements

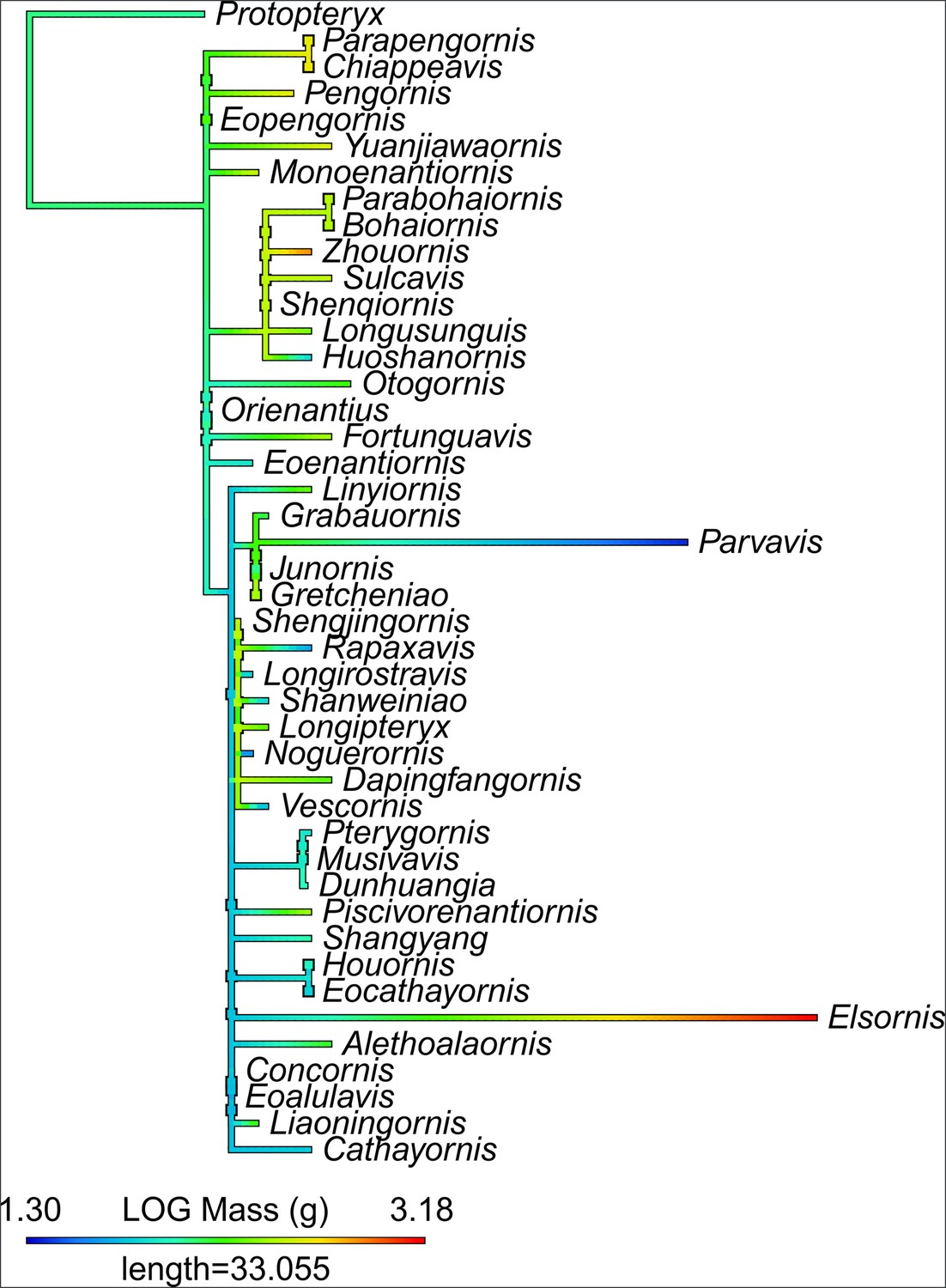
Ancestral state reconstruction of enantiornithine diet.
Phylogeny is presented as not time-scaled for node visibility. All enantiornithine taxa with dietary hypotheses are included, as well as enantiornithines complete enough to create robust mass estimates (Miller and Pittman, 2021; Miller et al., 2022; Miller et al., 2023; Serrano et al., 2015) and the non-ornithothoracine pygostylian Sapeornis as an outgroup. Taxa with bold names have diet assigned based on preserved meals (Eoalulavis [Sanz et al., 1996], Sapeornis [O’Connor, 2019a]) or quantitative diet proxies (Miller et al., 2022; Miller et al., 2023). Remaining diets assignments are based on qualitative morphology and depositional setting (Table 11). The diet of the common ancestor of Enantiornithes remains obscure, though many late-diverging enantiornithines are recovered as ancestrally invertivorous.
Figure 9—figure supplement 1
Ancestral state reconstruction of enantiornithine body mass.
Sapeornis is included as an outgroup. Mass data is taken from Table 3 and past work (Miller et al., 2022; Miller et al., 2023; Serrano et al., 2015). Tree is time-scaled and resolves polytomies in the order taxa appear on the tree. This does not account for other possible permutations of the tree topology; a more valid mass estimate for the common ancestor of Enantiornithes is provided in Table 7.
Figure 9—figure supplement 2
Ancestral state reconstruction of enantiornithine mechanical advantage (MA) and functional indices.
Sapeornis is included as an outgroup. AMA (A, B), PMA (C, D), OMA (E, F), AO (G, H), MCH (I), MMH (J), ACH (K), and AMH (L) reconstructions are provided for the upper (A, C, E, G, I, K) and lower (B, D, F, H, J, L) jaws. Character abbreviations: AMA, anterior jaw-closing mechanical advantage; PMA, posterior jaw-closing mechanical advantage; OMA, jaw-opening mechanical advantage; AO, relative articular offset; MCH, relative maximum cranial height; MMH, relative maximum mandible height; ACH, relative average cranial height; AMH, relative average mandible height. Suffixes of upr and lwr respectively denote the measurement is taken of the upper or lower jaw. Enantiornithine data is taken from this study and past work (Miller et al., 2022; Miller et al., 2023); the outgroup Sapeornis is measured from the reconstruction in Hu et al., 2020a. Tree is time-scaled and resolves polytomies in the order taxa appear on the tree. This does not account for other possible permutations of the tree topology; more valid estimates of variates for the common ancestor of Enantiornithes are provided in Table 7.
Figure 9—figure supplement 3
Ancestral state reconstruction of enantiornithine mesh-weighted arithmetic mean (MWAM) strain under bite loading in finite element analysis (FEA).
Sapeornis is included as an outgroup. Enantiornithine data is taken from this study and past work (Miller et al., 2022; Miller et al., 2023); data from Sapeornis is taken from a model constructed by Yuen Ting (Athena) Tse for an in-progress collaborative work based on the skull reconstruction in Hu et al., 2020a. Tree is time-scaled and resolves polytomies in the order taxa appear on the tree. This does not account for other possible permutations of the tree topology; a more valid MWAM strain estimate for the common ancestor of Enantiornithes is provided in Table 7.
Figure 9—figure supplement 4
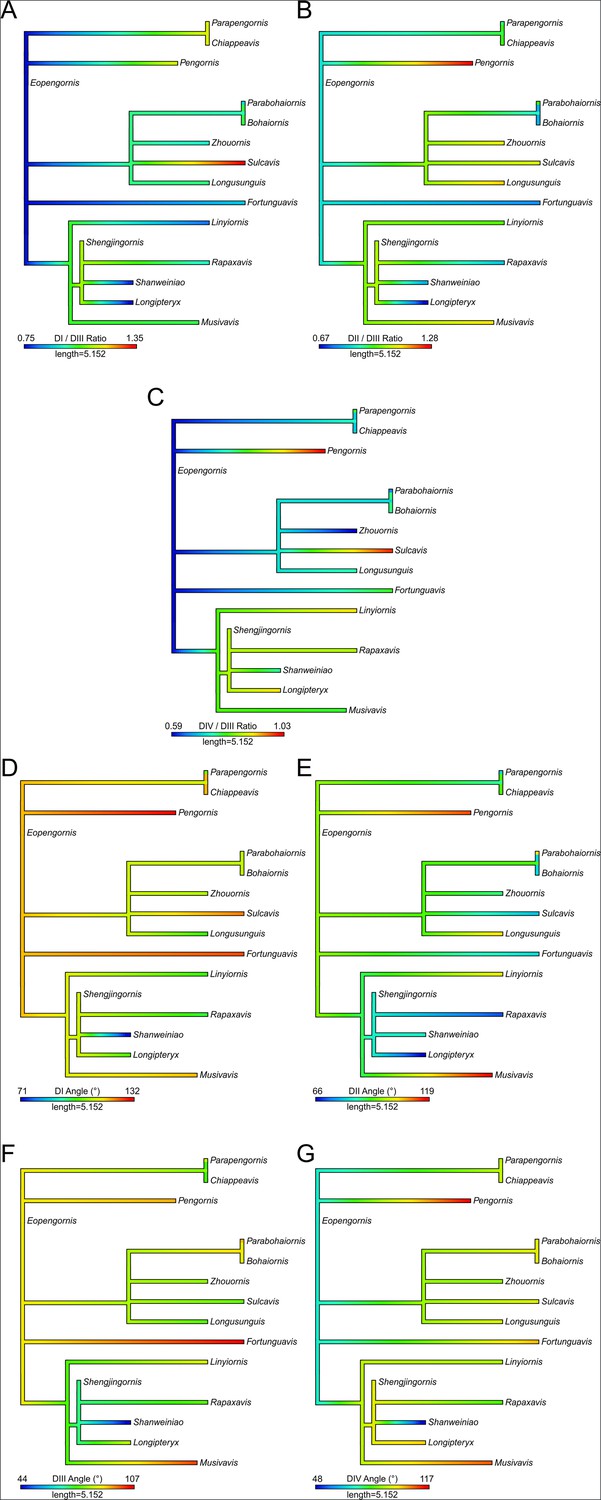
Ancestral state reconstruction of enantiornithine pedal traditional morphometric (TM) variables.
Sapeornis is included as an outgroup. Reconstructions of ratios of DI (A), DII (B), and DIV (C) arc length relative to DIII arc length as well as DI (D), DII (E), DIII (F), and DIV (G) angles are provided. Data is taken from this study and past work (Miller et al., 2022; Miller et al., 2023; Pittman et al., 2022). Tree is time-scaled and resolves polytomies in the order taxa appear on the tree. This does not account for other possible permutations of the tree topology; more valid estimates of variates for the common ancestor of Enantiornithes are provided in Table 7.
Figure 10
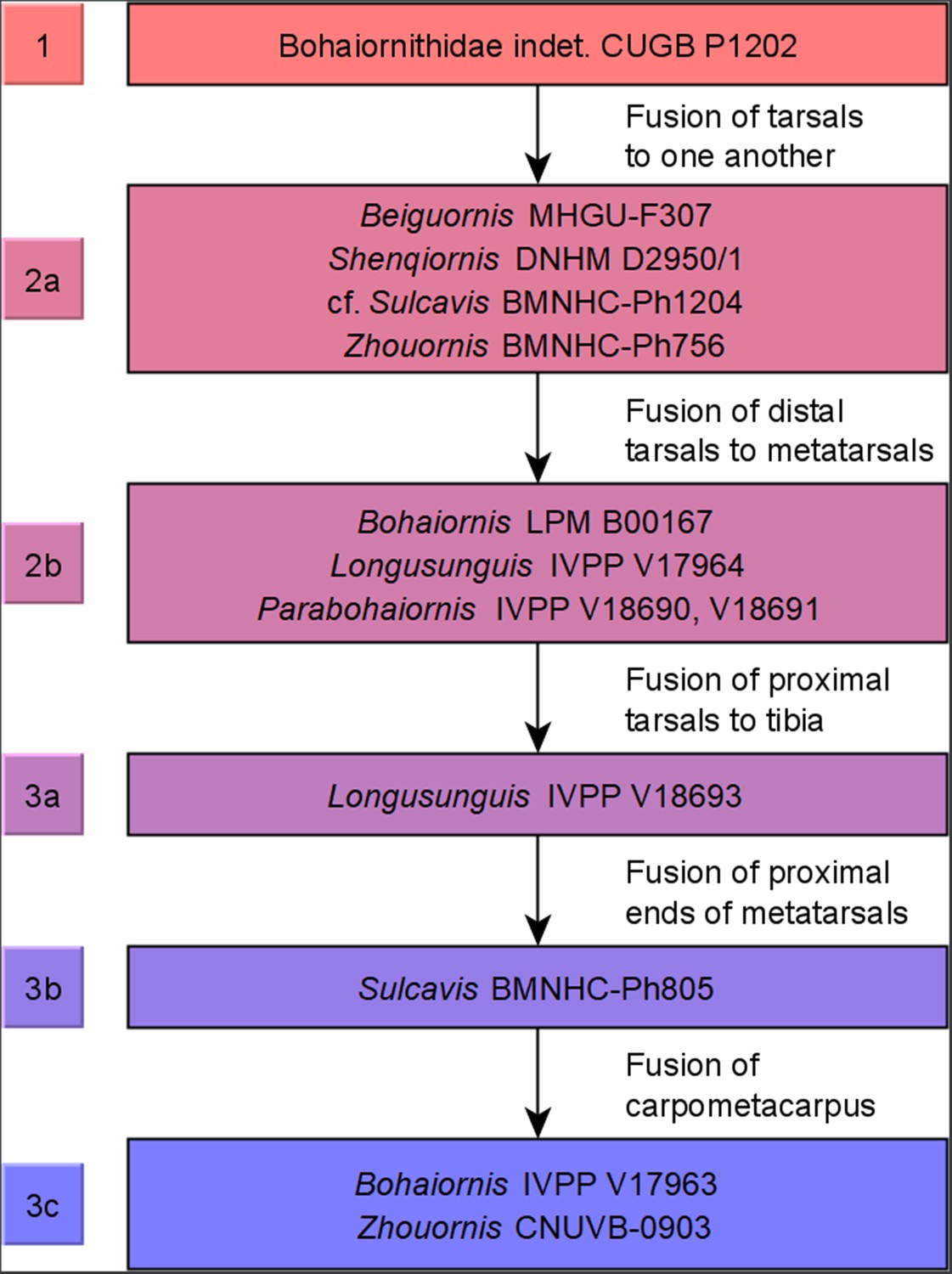
Ontogenetic stages of bohaiornithids.
Stages are based on Hu and O’Connor, 2017, with new subdivisions of stage 3 (possibly specific to Bohaiornithidae) as noted in ‘Methods’. Subadult status is reached at or before stage 1 and adulthood within stage 3; see Table 10 for details.
Tables
Table 1
Summary of taxa included in Bohaiornithidae.
‘Bohaiornithidae’ is used in an informal term to refer to any distinct clade containing Bohaiornis, as the strict clade definition is unstable (Liu et al., 2022). If multiple members of the other five originally defined bohaiornithids (Wang et al., 2014b) resolved near Bohaiornis, ‘Bohaiornithidae’ was considered to be the least inclusive clade containing all of them with a tolerance of two bird taxa not in the original six between any two internal nodes.
| Status | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| ‘Bohaiornithidae’ | + | |||||||||||
| Sister to ‘Bohaiornithidae’ | ~ | |||||||||||
| Not ‘Bohaiornithidae’ | - | |||||||||||
| Not in study | x | |||||||||||
| Beiguornis | Bohaiornis | Longusunguis | Parabohaiornis | Shenqiornis | Sulcavis | Zhouornis | Gretcheniao | Musivavis | Eoenantiornis | Fortunguavis | Linyiornis | |
| Wang, 2014a | x | + | + | + | + | + | + | x | x | ~ | x | x |
| Wang et al., 2015c | x | + | + | + | + | + | + | x | x | - | - | x |
| Wang et al., 2015a | x | + | + | + | + | + | + | x | x | ~ | - | x |
| Wang and Liu, 2016a | x | + | + | + | + | + | + | x | x | ~ | ~ | x |
| Wang et al., 2016b | x | + | + | + | + | + | + | x | x | ~ | + | + |
| Hu and O’Connor, 2017 | x | + | + | + | + | + | + | x | x | - | + | x |
| Wang and Zhou, 2017b | x | + | + | + | + | + | + | x | x | - | ~ | ~ |
| Cau, 2018 | x | + | x | x | x | + | + | x | x | x | x | x |
| Chiappe et al., 2019 | x | + | + | + | - | - | - | - | x | - | - | x |
| Zhang and Wang, 2019 | x | + | + | + | + | + | + | x | x | x | ~ | ~ |
| Hu et al., 2020b | x | + | + | + | + | + | - | x | x | - | - | - |
| O’Connor et al., 2020b | x | + | ~ | + | ~ | ~ | ~ | x | x | ~ | ~ | ~ |
| Pittman, 2020b, new technology | x | + | + | + | + | + | + | x | x | - | - | - |
| Pittman, 2020b, traditional | x | + | + | + | + | + | + | x | x | ~ | ~ | ~ |
| Wang and Zhou, 2020b | x | + | + | + | + | + | + | x | x | - | - | - |
| Li et al., 2022, strict consensus | x | + | ~ | + | ~ | ~ | ~ | x | x | ~ | x | x |
| Li et al., 2022, reduced consensus | x | + | + | + | + | + | + | x | x | ~ | x | x |
| Liu et al., 2022 | x | + | + | + | - | + | + | ~ | x | - | - | x |
| Wang et al., 2021b | x | + | + | + | + | + | + | - | x | - | - | - |
| Wang, 2022a | + | + | - | + | + | + | + | - | x | + | + | x |
| Wang et al., 2022b, unweighted | x | + | - | x | + | + | + | - | - | + | + | x |
| Wang et al., 2022b, K = 20 | x | + | - | - | + | - | - | - | - | - | - | x |
| Wang et al., 2022b, K = 5 | x | + | - | - | + | - | + | - | - | + | - | x |
Table 2
Diet cut-offs used in this study.
Percentages refer to values given in EltonTraits 1.0 (Wilman et al., 2014), with Diet-Tetr being the sum of Diet-Ect and Diet-End (ectotherm and endotherm tetrapod food sources are combined). Granivores were separated into husking and swallowing subdivisions based on feeding descriptions in the literature.
| Diet | Cut-off |
|---|---|
| Folivore | 60+% Diet-PlantO |
| Frugivore | 60+% Diet-Fruit |
| Generalist | 40% or less in any category |
| Granivore | 70+% Diet-Seed |
| Invertivore | 60+% Diet-Inv |
| Nectarivore | 60+% Diet-Nect |
| Piscivore | 50+% Diet-Fish |
| Scavenger | 50+% Diet-Scav |
| Tetrapod Hunter | 60+% Diet-Tetr |
Table 3
Masses for bohaiornithid taxa based on the regression equations of Serrano et al., 2015.
Most masses were previously reported in Miller and Pittman, 2021, though masses for the juvenile bohaiornithid CUGB P1202 (Peteya et al., 2017), Beiguornis khinganensis MHGU-F307/8 (Wang, 2022a), and cf. Sulcavis BMNHC-Ph1204 (Liu et al., 2022) are newly calculated in this study from literature images.
| Taxon | Specimen | Ontogenetic stage | Mean mass estimate (g) | Min mass estimate (g) | Max Mass Estimate (g) |
|---|---|---|---|---|---|
| Bohaiornithidae indet. | CUGB P1202 | 1 | 113 | 91 | 135 |
| Beiguornis khinganensis | MHGU-F307/8 | 2a | 260 | 210 | 310 |
| Bohaiornis guoi | IVPP V17963 | 3c | 300 | 242 | 358 |
| B. guoi | LPM B00167 | 2b | 249 | 201 | 298 |
| Longusunguis kurochkini | IVPP V17964 | 2b | 171 | 137 | 204 |
| L. kurochkini | IVPP V18693 | 3a | 237 | 191 | 283 |
| Parabohaiornis martini | IVPP V18691 | 2b | 221 | 178 | 263 |
| Parabohaiornis martini | IVPP V28398 | 3b | 323 | 260 | 386 |
| Shenqiornis mengi | DNHM D2950/1 | 2a | 340 | 274 | 406 |
| Sulcavis geeorum | BMNHC-Ph805 | 3b | 333 | 268 | 397 |
| cf. Sulcavis | BMNHC-Ph1204 | 2a | 171 | 137 | 204 |
| Zhouornis hani | BMNHC-Ph756 | 2a | 253 | 204 | 303 |
| Z. hani | CNUVB-903 | 3c | 758 | 611 | 905 |
Table 4
Posterior probabilities predicting bohaiornithid diet by flexible discriminate analysis (FDA) from mechanical advantage (MA) and functional indices of extant bird jaws.
Values in blue are most likely, values in red are least likely. All bohaiornithids have high affinity with generalists and low affinity with husking granivores, with other affinities varying by taxon. Diet abbreviations: GranivoreH, Husking Granivore; GranivoreS, Swallowing Granivore; Tetra Hunt, Tetrapod Hunter.
| Taxon | Folivore | Frugivore | Generalist | GranivoreH | GranivoreS | Invertivore | Nectarivore | Piscivore | Scavenger | Tetra Hunt |
|---|---|---|---|---|---|---|---|---|---|---|
| Bohaiornis | 9.99E-01 | 6.43E-06 | 2.92E-04 | 5.97E-13 | 2.15E-04 | 5.43E-06 | 1.62E-05 | 2.00E-07 | 1.74E-10 | 7.78E-08 |
| Longusunguis | 1.53E-02 | 1.81E-01 | 5.39E-01 | 5.48E-07 | 4.38E-03 | 1.57E-01 | 3.05E-02 | 6.91E-02 | 9.85E-05 | 3.95E-03 |
| Parabohaiornis | 2.44E-01 | 6.72E-04 | 2.34E-01 | 2.49E-07 | 3.64E-01 | 1.05E-02 | 2.42E-04 | 1.49E-02 | 1.32E-01 | 7.69E-05 |
| Shenqiornis | 7.32E-03 | 1.12E-02 | 6.45E-01 | 1.81E-06 | 2.00E-03 | 5.15E-02 | 8.12E-03 | 2.65E-01 | 9.50E-03 | 5.24E-04 |
| Sulcavis | 7.28E-02 | 1.58E-02 | 8.22E-01 | 5.46E-07 | 4.12E-03 | 2.43E-02 | 3.39E-03 | 5.68E-02 | 5.74E-04 | 6.25E-04 |
| Zhouornis | 3.20E-02 | 1.61E-01 | 3.11E-01 | 9.82E-07 | 2.30E-02 | 2.69E-01 | 5.42E-02 | 7.86E-02 | 9.64E-04 | 6.99E-02 |
Table 5
Posterior probabilities predicting bohaiornithid diet by flexible discriminate analysis (FDA) from finite element analysis (FEA) following the intervals method (Marcé-Nogué et al., 2017).
Values in blue are most likely, values in red are least likely. Bohaiornithid affinities varying considerably between taxa, only universally not resembling tetrapod hunters. Diet abbreviations: GranivoreH, Husking Granivore; GranivoreS, Swallowing Granivore; Tetra Hunt, Tetrapod Hunter.
| Taxon | Folivore | Frugivore | Generalist | GranivoreH | GranivoreS | Invertivore | Nectarivore | Piscivore | Scavenger | Tetra Hunt |
|---|---|---|---|---|---|---|---|---|---|---|
| Bohaiornis | 3.44E-10 | 9.33E-12 | 1.73E-07 | 2.25E-23 | 1.00E+00 | 7.09E-07 | 4.67E-19 | 1.43E-10 | 8.82E-06 | 3.37E-19 |
| Longusunguis | 3.05E-01 | 1.15E-13 | 6.95E-01 | 1.56E-26 | 2.26E-15 | 3.58E-09 | 4.35E-26 | 1.51E-11 | 3.47E-29 | 1.07E-20 |
| Parabohaiornis | 8.49E-02 | 3.30E-15 | 2.08E-13 | 3.50E-24 | 5.52E-01 | 1.18E-04 | 4.67E-04 | 3.63E-01 | 3.12E-13 | 1.13E-06 |
| Shenqiornis | 2.28E-08 | 1.45E-23 | 9.39E-09 | 4.31E-11 | 2.28E-13 | 2.47E-05 | 3.61E-02 | 9.58E-01 | 6.11E-03 | 2.81E-20 |
| Sulcavis | 6.82E-06 | 1.91E-16 | 1.19E-03 | 9.94E-01 | 5.99E-18 | 5.65E-09 | 4.71E-03 | 1.22E-07 | 4.68E-35 | 1.55E-22 |
| Zhouornis | 5.69E-50 | 1.00E+00 | 5.55E-20 | 4.89E-13 | 4.06E-12 | 2.08E-18 | 8.99E-13 | 4.74E-20 | 1.13E-53 | 2.52E-17 |
Table 6
Posterior probabilities predicting bohaiornithid pedal ecology by flexible discriminate analysis (FDA) and phylogenetic flexible discriminate analysis (pFDA) from traditional morphometric (TM) of extant bird claws.
Values in blue are most likely, values in red are least likely. Bohaiornithid affinities vary considerably by taxon and between FDA and pFDA. Diet abbreviations: GranivoreH, Husking Granivore; GranivoreS, Swallowing Granivore; Tetra Hunt, Tetrapod Hunter. Category abbreviations: large raptor, raptor taking prey which does not fit in the foot; small raptor, raptor taking prey which can fit in the foot.
| Test | Taxon | Ground | Perch | Large raptor | Small raptor | Shrike |
|---|---|---|---|---|---|---|
| FDA | Bohaiornis LPM B00167 | 2.45E-02 | 4.03E-01 | 2.69E-01 | 3.03E-01 | 4.60E-04 |
| Bohaiornis IVPP V17963 | 1.92E-02 | 2.52E-01 | 2.31E-04 | 7.23E-01 | 5.78E-03 | |
| Longusunguis | 3.65E-02 | 4.93E-01 | 3.76E-01 | 9.45E-02 | 4.42E-07 | |
| Parabohaiornis IVPP V18690 | 2.94E-05 | 8.57E-01 | 1.49E-03 | 5.57E-02 | 8.63E-02 | |
| Parabohaiornis IVPP V18691 | 2.41E-05 | 9.66E-01 | 2.66E-02 | 4.41E-03 | 2.85E-03 | |
| Sulcavis | 2.03E-01 | 1.80E-02 | 2.17E-01 | 5.62E-01 | 2.93E-09 | |
| Zhouornis CNUVB-0903 | 1.22E-01 | 3.62E-01 | 6.10E-02 | 4.07E-01 | 4.83E-02 | |
| Zhouornis BMNHC Ph 756 | 5.46E-01 | 1.41E-02 | 1.72E-02 | 4.22E-01 | 1.33E-04 | |
| pFDA | Bohaiornis LPM B00167 | 0.00E+00 | 6.09E-02 | 1.29E-01 | 8.10E-01 | 4.57E-05 |
| Bohaiornis IVPP V17963 | 0.00E+00 | 6.21E-07 | 4.58E-06 | 4.87E-12 | 1.00E+00 | |
| Longusunguis | 0.00E+00 | 2.66E-03 | 9.94E-01 | 2.82E-03 | 6.46E-04 | |
| Parabohaiornis IVPP V18690 | 0.00E+00 | 1.98E-04 | 2.10E-01 | 7.90E-01 | 3.27E-12 | |
| Parabohaiornis IVPP V18691 | 0.00E+00 | 2.68E-02 | 3.05E-01 | 6.68E-01 | 3.28E-07 | |
| Sulcavis | 0.00E+00 | 2.87E-04 | 3.92E-05 | 3.27E-05 | 1.00E+00 | |
| Zhouornis CNUVB-0903 | 0.00E+00 | 4.94E-01 | 2.84E-01 | 2.19E-01 | 3.00E-03 | |
| Zhouornis BMNHC Ph 756 | 0.00E+00 | 3.10E-01 | 5.23E-03 | 3.96E-03 | 6.80E-01 |
Table 7
Quantitative ancestral states of the common ancestor of Enantiornithines.
Mean values are the average of 10,000 random tree permutations where polytomies are randomly resolved. 95% confidence intervals are given as the 2.5 and 97.5% quantiles of the permutations, though upper and lower bounds for all values are identical to seven significant figures. As these values are based on multiple possible topologies of the enantiornithine tree, we consider them more valid than those visualised in figure supplements of Figure 9.
| Line of evidence | Trait | Mean | 95% CI low | 95% CI high |
|---|---|---|---|---|
| Mass | Body mass (g) | 198 | 198 | 198 |
| MA | AMAup | 0.13 | 0.13 | 0.13 |
| PMAup | 0.19 | 0.19 | 0.19 | |
| OMAup | 0.19 | 0.19 | 0.19 | |
| AOup | 0.09 | 0.09 | 0.09 | |
| MCHup | 0.32 | 0.32 | 0.32 | |
| ACHup | 0.19 | 0.19 | 0.19 | |
| AMAlow | 0.21 | 0.21 | 0.21 | |
| PMAlow | 0.32 | 0.32 | 0.32 | |
| OMAlow | 0.08 | 0.08 | 0.08 | |
| AOlow | 0.065 | 0.065 | 0.065 | |
| MMHlow | 0.12 | 0.12 | 0.12 | |
| AMHlow | 0.07 | 0.07 | 0.07 | |
| FEA | MWAM strain (µε) | 227 | 227 | 227 |
| Pedal TM | DI/DIII ratio | 0.76 | 0.76 | 0.76 |
| DII/DIII ratio | 0.85 | 0.85 | 0.85 | |
| DIV/DIII ratio | 0.60 | 0.60 | 0.60 | |
| DI angle (°) | 120 | 120 | 120 | |
| DII angle (°) | 98 | 98 | 98 | |
| DIII angle (°) | 90 | 90 | 90 | |
| DIV angle (°) | 71 | 71 | 71 |
-
MA = mechanical advantage; FEA = finite element analysis; TM = traditional morphometric; AMA = anterior jaw-closing mechanical advantage; PMA = posterior jaw-closing mechanical advantage; OMA = jaw-opening mechanical advantage; AO = relative articular offset; MCH = relative maximum cranial height; MMH = relative maximum mandible height; ACH = relative average cranial height; AMH = relative average mandible height; MWAM = mesh-weighted arithmetic mean.
Table 8
Posterior probability of diet in the common ancestor of Enantiornithes based on mechanical advantage (MA) and functional indices.
Generalist feeding is the most likely, followed by piscivory and invertivory. Diet abbreviations: GranivoreH, Husking Granivore; GranivoreS, Swallowing Granivore; Tetra Hunt, Tetrapod Hunter.
| Folivore | Frugivore | Generalist | GranivoreH | GranivoreS | Invertivore | Nectarivore | Piscivore | Scavenger | Tetra Hunt |
|---|---|---|---|---|---|---|---|---|---|
| 2.53E-03 | 5.58E-03 | 6.51E-01 | 2.17E-07 | 6.44E-03 | 7.57E-02 | 3.99E-03 | 2.12E-01 | 4.28E-02 | 4.41E-04 |
Table 9
Summary table of interpretations of each line of evidence used herein.
Body mass, mechanical advantage (MA), and finite element analysis (FEA) inform diet. Pedal traditional morphometric (TM) informs the use of the pes or lack thereof in feeding. See relevant discussion sections for additional details. Bolded diets are agreed upon by all available diet proxies. Bolded pedal ecologies are either help discriminate diet (carnivory in Longusunguis) or are supported over other possibilities by diet information (non-raptorial perching in Bohaiornis). Given the uncertain application of skull TM to bohaiornithids, diets from this proxy are not bolded.
| Line of evidence | Taxon | Likely diets/ecologies | Unlikely diets/ecologies |
|---|---|---|---|
| Body mass | Beiguornis | Folivore, Frugivore, Generalist, Piscivore, Tetrapod Hunter | Granivore, Invertivore, Nectarivore |
| Bohaiornis | Folivore, Frugivore, Generalist, Piscivore, Tetrapod Hunter | Granivore, Invertivore, Nectarivore | |
| Longusunguis | Folivore, Frugivore, Generalist, Piscivore, Tetrapod Hunter | Granivore, Invertivore, Nectarivore | |
| Parabohaiornis | Folivore, Frugivore, Generalist, Piscivore, Tetrapod Hunter | Granivore, Invertivore, Nectarivore | |
| Shenqiornis | Folivore, Frugivore, Generalist, Piscivore, Tetrapod Hunter | Granivore, Invertivore, Nectarivore | |
| Sulcavis | Folivore, Frugivore, Generalist, Piscivore, Tetrapod Hunter | Granivore, Invertivore, Nectarivore | |
| Zhouornis | Folivore, Frugivore, Generalist, Piscivore, Scavenger, Tetrapod Hunter | Granivore, Invertivore, Nectarivore | |
| Mechanical advantage | Bohaiornis | Folivore, Generalist, Swallowing Granivore | Husking Granivore, Scavenger |
| Longusunguis | Frugivore, Generalist, Invertivore, Piscivore | Husking Granivore, Scavenger | |
| Parabohaiornis | Folivore, Generalist, Scavenger, Swallowing Granivore | Husking Granivore, Tetrapod Hunter | |
| Shenqiornis | Folivore, Frugivore, Generalist, Invertivore, Piscivore | Husking Granivore, Tetrapod Hunter | |
| Sulcavis | Folivore, Generalist, Invertivore, Piscivore | Husking Granivore, Scavenger | |
| Zhouornis | Frugivore, Generalist, Invertivore, Piscivore | Husking Granivore, Scavenger | |
| Finite element analysis | Bohaiornis | Folivore, Husking Granivore, Nectarivore | Swallowing Granivore, Invertivore, Piscivore, Scavenger, Tetrapod Hunter |
| Longusunguis | Folivore, Frugivore, Generalist, Husking Granivore, Swallowing Granivore, Invertivore, Nectarivore, Piscivore | Scavenger, Tetrapod Hunter | |
| Parabohaiornis | Folivore, Frugivore, Generalist, Husking Granivore, Swallowing Granivore, Invertivore, Nectarivore, Piscivore | Scavenger, Tetrapod Hunter | |
| Shenqiornis | Folivore, Frugivore, Generalist, Husking Granivore, Invertivore, Nectarivore, Piscivore | Swallowing Granivore, Scavenger, Tetrapod Hunter | |
| Sulcavis | Folivore, Frugivore, Generalist, Husking Granivore, Invertivore, Nectarivore, Piscivore | Swallowing Granivore, Scavenger, Tetrapod Hunter | |
| Zhouornis | Frugivore, Generalist, Husking Granivore, Nectarivore | Swallowing Granivore, Invertivore, Piscivore, Scavenger, Tetrapod Hunter | |
| Pedal traditional morphometrics | Bohaiornis | Non-Raptorial Perching, Small Raptor | Ground Bird, Shrike-like |
| Longusunguis | Large Raptor | Ground Bird, Shrike-like | |
| Parabohaiornis | Non-Raptorial Perching | Ground Bird | |
| Sulcavis | Large Raptor, unique ecology not in dataset | Shrike-like | |
| Zhouornis | Small Raptor, Non-Raptorial Perching | Ground Bird, Shrike-like | |
| Skull traditional morphometrics | Bohaiornis | Folivore, Frugivore, Generalist | Granivore, Nectarivore |
| Longusunguis | Folivore, Frugivore, Generalist | Granivore, Nectarivore | |
| Parabohaiornis | Folivore, Frugivore, Generalist | Granivore, Nectarivore | |
| Shenqiornis | Unique ecology not in dataset | All sampled diets | |
| Sulcavis | Unique ecology not in dataset | All sampled diets | |
| Zhouornis | Unique ecology not in dataset | All sampled diets |
Table 10
Comparison of the bone character stages of Hu and O’Connor, 2017 with the histological maturity stages of Atterholt et al., 2021.
Subadult maturity appears to be obtained by stage 1 (the most immature fusion stage of any specimen examined in this work), while full skeletal maturity is not obtained until after stage 3 (the most mature fusion stage). Histological maturities are identified by Atterholt et al., 2021 directly, bone fusion stages are taken from Hu and O’Connor, 2017 where possible and intuited based on their fusion criteria where not.
| Specimen | Atterholt et al.Histological maturity | Hu and O'ConnorCharacter stage |
|---|---|---|
| Parapengornis IVPP V18687 | Young subadult | 1 |
| Eopengornis STM24-1 | Young subadult | 2a |
| Monoenantiornis IVPP V20289 | Subadult | 2a |
| Parvavis IVPP V18586 | Subadult | 2b |
| Cruralispennia IVPP V21711 | Subadult | 3 |
| Pterygornis IVPP V16363 | Young adult | 3 |
| Avimaia IVPP V25371 | Adult | 3 |
| Mirarce UCMP139500 | Adult | 3 |
| Mirusavis IVPP V18692 | Adult | 3 |
| STM 29-8 | Adult | 3 |
| Zhouornis CNUVB-0908 | Adult | 3 |
Table 11
Qualitative dietary hypotheses for enantiornithine taxa.
The hypothesised diet listed may overgeneralise the hypothesis of the original publication for the sake of brevity. Most notably we often simplify hypotheses like ‘carnivory taking small animals’ to invertivory, as when these authors provide analogous extant birds for context, they predominantly were invertivorous birds.
Additional files
-
MDAR checklist
- https://cdn.elifesciences.org/articles/89871/elife-89871-mdarchecklist1-v1.pdf
-
Supplementary file 1
K and Kmult values for each extant dataset.
- https://cdn.elifesciences.org/articles/89871/elife-89871-supp1-v1.xlsx
-
Supplementary file 2
K values for individual MA and functional index variables of the extant skull dataset.
- https://cdn.elifesciences.org/articles/89871/elife-89871-supp2-v1.xlsx
-
Supplementary file 3
Significant differences between extant diet groups, based on phylogenetic HSD of MA and functional indices.
- https://cdn.elifesciences.org/articles/89871/elife-89871-supp3-v1.xlsx
-
Supplementary file 4
Sensitivity analysis of quadrate position affecting bohaiornithid predictions diet by FDA from MA and functional indices of extant bird jaws.
- https://cdn.elifesciences.org/articles/89871/elife-89871-supp4-v1.xlsx
-
Supplementary file 5
Significant differences between extant diet groups, based on phylogenetic HSD of FEA intervals data.
- https://cdn.elifesciences.org/articles/89871/elife-89871-supp5-v1.xlsx
-
Supplementary file 6
K values for individual variables used in pedal TM analyses.
- https://cdn.elifesciences.org/articles/89871/elife-89871-supp6-v1.xlsx
-
Supplementary file 7
Significant differences between extant pedal ecology groups, based on phylogenetic HSD of pedal TM data.
- https://cdn.elifesciences.org/articles/89871/elife-89871-supp7-v1.xlsx
-
Supplementary file 8
K values for individual variables used in skull TM analyses.
- https://cdn.elifesciences.org/articles/89871/elife-89871-supp8-v1.xlsx
Download links
A two-part list of links to download the article, or parts of the article, in various formats.
Downloads (link to download the article as PDF)
Open citations (links to open the citations from this article in various online reference manager services)
Cite this article (links to download the citations from this article in formats compatible with various reference manager tools)
Synthetic analysis of trophic diversity and evolution in Enantiornithes with new insights from Bohaiornithidae
eLife 12:RP89871.
https://doi.org/10.7554/eLife.89871.3




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}