The acid ceramidase/ceramide axis controls parasitemia in Plasmodium yoelii-infected mice by regulating erythropoiesis
- Institute of Medical Microbiology, University Hospital Essen, University Duisburg-Essen, Germany
- Institute of Pharmacy, Freie Universität Berlin, Germany
- Molecular Parasitology, Institute of Biology/Faculty for Life Sciences, Humboldt University Berlin, Germany
- Institute of Immunology, University Hospital Essen, University Duisburg-Essen, Germany
- Institute of Molecular Biology, University of Duisburg-Essen, Germany
Abstract
Acid ceramidase (Ac) is part of the sphingolipid metabolism and responsible for the degradation of ceramide. As bioactive molecule, ceramide is involved in the regulation of many cellular processes. However, the impact of cell-intrinsic Ac activity and ceramide on the course of Plasmodium infection remains elusive. Here, we use Ac-deficient mice with ubiquitously increased ceramide levels to elucidate the role of endogenous Ac activity in a murine malaria model. Interestingly, ablation of Ac leads to alleviated parasitemia associated with decreased T cell responses in the early phase of Plasmodium yoelii infection. Mechanistically, we identified dysregulated erythropoiesis with reduced numbers of reticulocytes, the preferred host cells of P. yoelii, in Ac-deficient mice. Furthermore, we demonstrate that administration of the Ac inhibitor carmofur to wildtype mice has similar effects on P. yoelii infection and erythropoiesis. Notably, therapeutic carmofur treatment after manifestation of P. yoelii infection is efficient in reducing parasitemia. Hence, our results provide evidence for the involvement of Ac and ceramide in controlling P. yoelii infection by regulating red blood cell development.
Editor's evaluation
This study provides evidence that murine acid ceramidase (Ac) is required for normal erythropoiesis and the development of rodent malaria. The findings are of interest in understanding molecular processes involved in regulating erythropoiesis, as well as the potential to develop host-directed therapies for malarial parasites that target human reticulocytes.
https://doi.org/10.7554/eLife.77975.sa0Introduction
Malaria remains one of the most life-threatening infectious diseases in the world, with estimated 241 million cases and 627,000 related deaths in 2020 (World health organization, 2021). Malaria is caused by infections with the parasite Plasmodium that undergoes a complex life cycle within its hosts. In the mammalian host, plasmodia invade red blood cells (RBCs) for their asexual propagation. Depending on the species, the parasites favor different RBC maturation stages for their intraerythrocytic development. While the most lethal human malaria species, Plasmodium falciparum is able to infect both, mature erythrocytes and immature reticulocytes, the most widespread parasite Plasmodium vivax is restricted to invade the latter (Garnham, 1966; Kitchen, 1938; Chwatt, 1948). The formation of RBCs, known as erythropoiesis, primarily takes place in the bone marrow, where hematopoietic stem cells give rise to multipotent progenitors that stepwise differentiate into erythroblasts and reticulocytes (Moras et al., 2017). Early reticulocytes egress from the bone marrow and constitute 1–3% of circulating RBCs before they develop into mature erythrocytes (Chin-Yee et al., 1991). During erythropoiesis, the cells start to produce hemoglobin, lose nuclei and organelles, and induce plasma membrane remodeling. While the erythroid lineage marker Ter119 is induced in erythroblasts and subsequently persistently expressed, expression of the transferrin receptor 1 (CD71) gets lost during the transition from reticulocytes to mature erythrocytes (Chen et al., 2009).
Several metabolic processes are involved in the regulation of erythropoiesis and therefore might have an impact on Plasmodium infection. The best-known example of metabolites essential for erythropoiesis is circulating iron, which is almost exclusively absorbed by erythropoietic demand. Plasma concentration, tissue distribution, and absorption are under the control of hepcidin (Kautz and Nemeth, 2014). Upon Plasmodium blood stage infection, hepcidin expression in the liver is upregulated, which in turn modulates parasitemia, disease outcome, and superinfection (Howard et al., 2007; Wang et al., 2011; Portugal et al., 2011; Spottiswoode et al., 2014), fully supporting clinical studies that found associations of asymptomatic P. falciparum and P. vivax parasitemia with increased hepcidin concentrations and anemia (de Mast et al., 2010; Casals-Pascual et al., 2012). Additional essential metabolic processes include fatty acid oxidation to preserve renewal of hematopoietic stem cells, amino acid-induced mTOR signaling, and glutamine metabolism (Oburoglu et al., 2016). For terminal erythropoiesis, alterations in the lipid metabolism with reduced phosphatidylcholine and enhanced sphingomyelin have been proposed to play a critical role (Huang et al., 2018). However, the effects of these metabolic processes on Plasmodium blood infection remain elusive.
During the erythrocytic stage of malaria, CD4+ T cells play a crucial role in regulating the immune response. By secreting IFN-γ, they activate other immune cells, for example, macrophages, which are necessary for parasite clearance and control. Moreover, CD4+ T cells provide help for B cells to induce protective responses against the blood-stage parasites (Perez-Mazliah and Langhorne, 2014). Recently, we showed that modulations of the sphingolipid (SL) metabolism enhance T cell activity during Plasmodium yoelii infection (Hose et al., 2019). SLs are important components of biological membranes. Besides their importance in cell integrity, they have drawn attention as bioactive molecules, playing a crucial role in cell function and fate (Hannun and Obeid, 2018). The SL metabolism consists of a network of various enzymes that catalyze the formation and degradation of the different SLs. Ceramide represents a central hub in this SL network. It can either be formed by de novo synthesis or by metabolism of complex SLs, for example, sphingomyelin (Hannun and Obeid, 2008). Ceramidases hydrolyze ceramide to sphingosine, which can be further phosphorylated by sphingosine kinases, forming sphingosine-1-phosphate (S1P). So far, five different human ceramidases have been described according to their pH optimum: acid ceramidase (Ac, N-acylsphingosine amidohydrolase 1, ASAH1), neutral ceramidase (Nc, ASAH2), and alkaline ceramidase 1–3 (Acer1-3, ACER1-3), each having a murine counterpart (Mao and Obeid, 2008). Ac is localized in the lysosomes, where it preferentially hydrolyzes ceramides with medium-chain length (C12–C16) (Ferlinz et al., 2001; Momoi et al., 1982). Mutations in the ASAH1 gene can lead to two different rare diseases, the lysosomal storage disorder Farber disease (FD) and an epileptic disorder called spinal muscular atrophy with progressive myoclonic epilepsy (SMA-PME) (Yu et al., 2018). Ac and ceramide have also been implicated to play a role in several infectious diseases. While the pharmacological inhibition of Ac by ceranib-2 decreases measles virus replication in vitro (Grafen et al., 2019), genetic overexpression of Ac rescues cystic fibrosis mice from pulmonary Pseudomonas aeruginosa infections in vivo (Becker et al., 2021). Lang et al., 2020 recently showed that Ac expression in macrophages limits the propagation of herpes simplex virus type 1 and protects mice from severe course of infection. However, the role of host cell-intrinsic Ac activity and ceramide during Plasmodium infection remains elusive.
Here, using the nonlethal rodent Plasmodium strain P. yoelii 17XNL as murine malaria model, we evaluated how Ac ablation and pharmacological inhibition modulate the course of infection in mice. We demonstrate that increased ceramide levels in Ac-deficient mice or treatment with the Ac inhibitor carmofur significantly reduce parasitemia in P. yoelii-infected mice. Mechanistically, we identified reduced generation of reticulocytes, the target cells of P. yoelii, upon deletion or inhibition of Ac.
Results
Ac-deficient mice accumulate ceramide
Since the genetic ablation of Ac is embryonically lethal in mice, we made use of inducible Asah1/Rosa26creER/+ (Ac KO) mice, which received tamoxifen 8, 6, and 4 days prior to analysis or P. yoelii infection (Figure 1A; Eliyahu et al., 2007; Li et al., 2002). Tamoxifen-treated Asah1/Rosa26+/+ (Ac WT) littermates served as control. As depicted in Figure 1B, Ac mRNA expression was ubiquitously abolished in all analyzed tissues in Ac KO mice. Consequently, ceramide concentrations determined by mass spectrometry were significantly increased in spleens from Ac KO mice compared to Ac WT mice (Figure 1C).
Figure 1
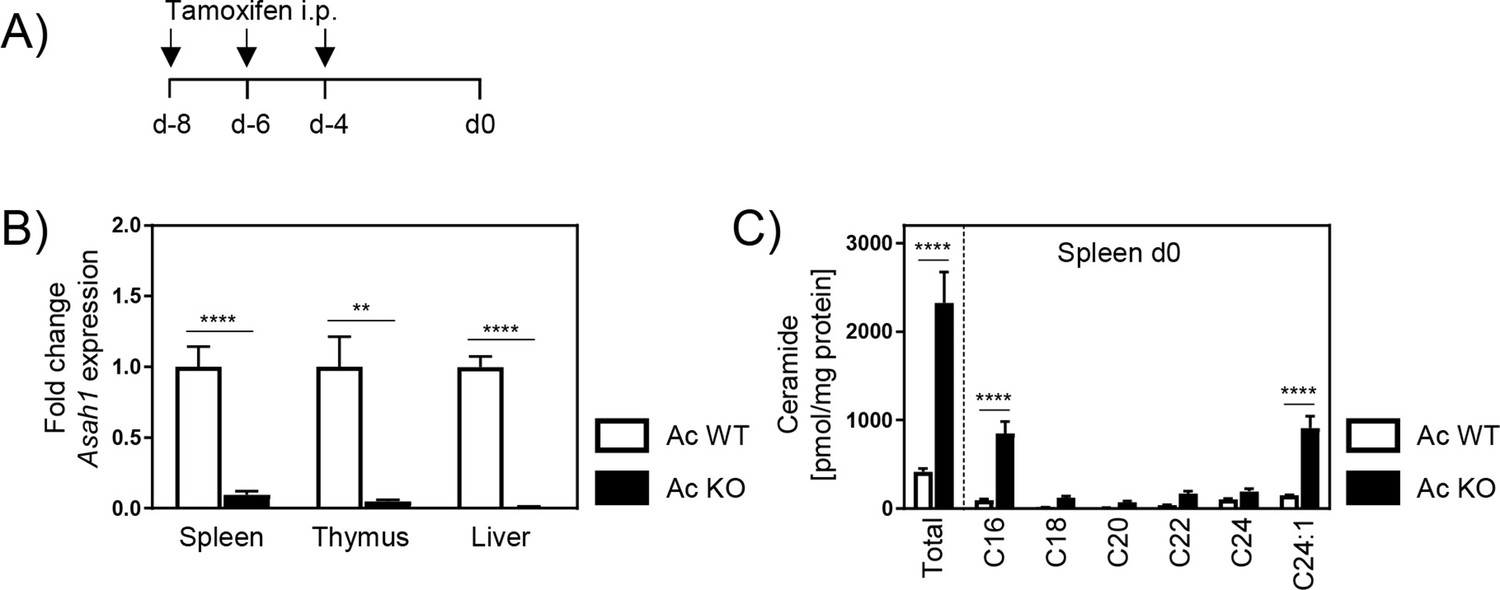
Ceramide accumulation in acid ceramidase (Ac) knockout (KO) mice.
(A) Induction of Ac deficiency in Asah1/Rosa26creER/+ mice (Ac KO) was achieved by intraperitoneal injection (i.p.) of 4 mg tamoxifen on days −8, –6, and –4. Tamoxifen-treated Asah1/Rosa26+/+ littermates (Ac WT) were used as control. (B) Ac KO validation was performed by analyzing Asah1 mRNA expression via RT-qPCR in spleen, thymus, and liver of Ac KO mice and Ac WT littermates as control on day 0 (n = 5–6). (C) Ceramide levels in spleen were determined by high-performance liquid chromatography mass spectrometry (HPLC-MS/MS) (n = 3–4). Data are presented as mean ± SEM. Statistical analyses were performed using Mann–Whitney U-test for (B) and two-way ANOVA, followed by Sidak’s post-test for (C) (**p<0.01, ****p<0.0001).
-
Figure 1—source data 1
Ceramide accumulation in acid ceramidase (Ac) knockout (KO) mice.
- https://cdn.elifesciences.org/articles/77975/elife-77975-fig1-data1-v2.xlsx
Ac deficiency results in reduced parasitemia and decreased T cell responses during early stage of P. yoelii infection
To study the role of Ac during malaria, we infected Ac KO mice and Ac WT littermates with P. yoelii. Interestingly, Ac KO mice showed significantly reduced parasitemia on days 3 and 7 post infection (p.i.) (Figure 2A). However, these differences were no longer detectable 10 and 14 days p.i. As splenomegaly is a common clinical signature of Plasmodium infections, we determined the spleen weight of infected Ac WT and Ac KO mice. Well in line with the observed effect on parasitemia, spleen weights of Ac KO mice were tremendously decreased compared to those of Ac WT mice 7 days p.i., but no longer differed on day 14 p.i. (Figure 2B).
Figure 2 with 1 supplement see all
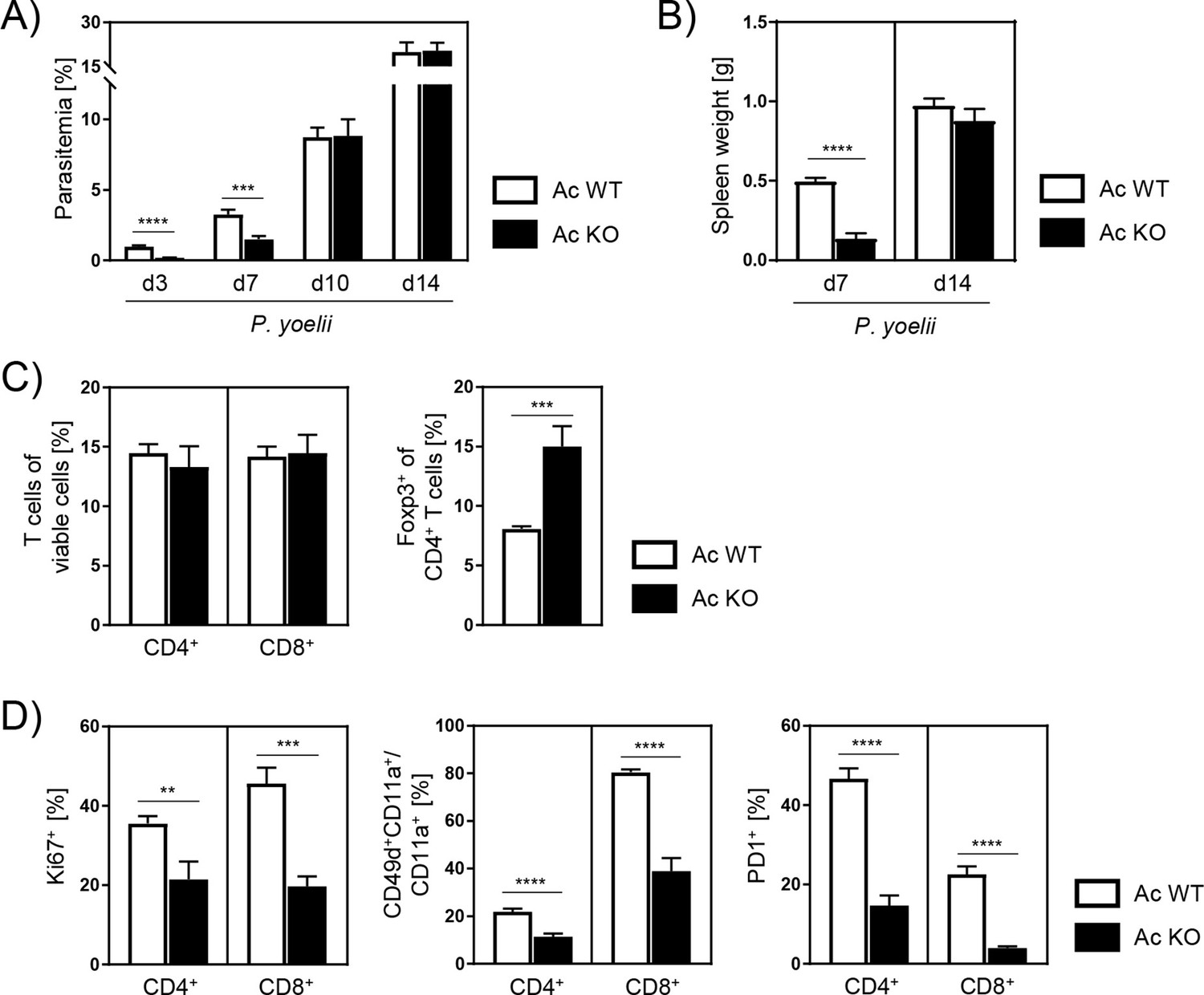
P.yoelii-infected acid ceramidase (Ac) knockout (KO) mice show decreased parasitemia with less T cell activation in the early phase of infection.
(A) Parasitemia of P. yoelii-infected Ac WT and Ac KO mice was determined at indicated time points by microscopy of Giemsa-stained blood films (n = 9–18). (B) Spleen weight on days 7 and 14 post infection (p.i.) (n = 7–9). (C) Frequencies of viable CD4+, CD8+, and regulatory T cells (Foxp3+ of CD4+) were analyzed by flow cytometry 7 days p.i. (n = 7–9). (D) Percentages of Ki67-, CD49dCD11a/CD11a-, and PD1-expressing CD4+ and CD8+ T cells were determined by flow cytometry 7 days p.i. (n = 7–9). Data from three to five independent experiments are presented as mean ± SEM. Statistical analyses were performed using Mann–Whitney U-test for (A) and unpaired Student’s t-test for (B–D) (**p<0.01, ***p<0.001, ****p<0.0001).
-
Figure 2—source data 1
P. yoelii-infected acid ceramidase (Ac) knockout (KO) mice show decreased parasitemia with less T cell activation in the early phase of infection.
- https://cdn.elifesciences.org/articles/77975/elife-77975-fig2-data1-v2.xlsx
Next, we analyzed the phenotype of T cells from spleen of infected mice 7 days p.i. by flow cytometry. T cells, especially CD4+ T cells, have been described to play an important role in limiting blood-stage malaria (Perez-Mazliah and Langhorne, 2014). While frequencies of CD4+ and CD8+ T cells did not differ between P. yoelii-infected Ac WT and Ac KO mice, we found significantly increased frequencies of CD4+Foxp3-expressing regulatory T cells (Tregs) in spleen of Ac KO mice compared to Ac WT littermates (Figure 2C). Interestingly, CD4+ and CD8+ T cells from Ac KO mice showed decreased expression of marker molecules associated with T cell activation, namely, Ki67, CD49d, CD11a, and PD1 (Figure 2D). In accordance with the observations regarding parasitemia and spleen weight, differences were diminished 14 days p.i. (Figure 2—figure supplement 1). These results show that Ac deficiency leads to reduced parasitemia in the early phase of P. yoelii infection accompanied by diminished T cell responses.
Cell-specific Ac deletion in T cells and myeloid cells does not affect the course of P. yoelii infection
In order to further evaluate the effect of Ac on immune responses during P. yoelii infection, we made use of cell type-specific Ac-deficient mice. Since T cells from infected global Ac KO mice were less activated, we next analyzed Asah1/Cd4cre mice that specifically lack Ac expression in CD4+ and CD8+ T cells (Figure 3A). However, T cell-specific Ac ablation had no impact on the course of P. yoelii infection as Ac CD4cre KO (Asah1/Cd4cre/+) and Ac CD4cre WT (Asah1/Cd4+/+) mice showed similar spleen weights and parasitemia 7 and 14 days p.i. (Figure 3B). We were wondering whether an early enhanced innate immune response as first line of defense contribute to an improved parasite clearance in global Ac KO mice and therefore to less T cell activation in the initial phase of infection. Thus, we used Asah1/Lyz2cre mice, which exhibit decreased Ac expression in myeloid cells, for example, macrophages, dendritic cells, and neutrophils (Figure 3C). However, upon P. yoelii infection, Ac Lyz2cre KO (Asah1/Lyz2cre/+) mice and Ac Lyz2cre WT (Asah1/Lyz2+/+) littermates did not significantly differ regarding parasitemia and spleen weight (Figure 3D).
Figure 3
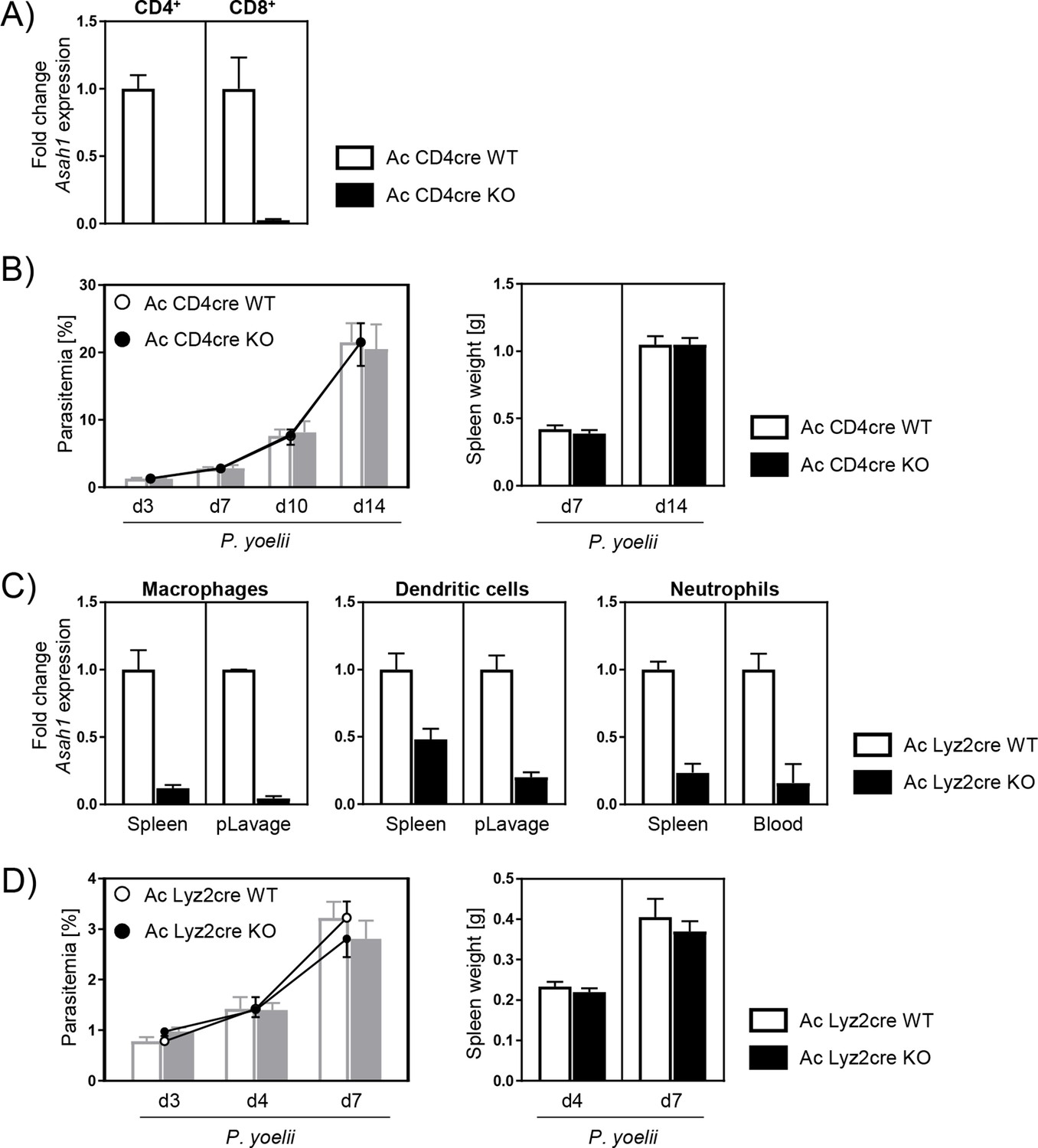
T cell-specific and myeloid-specific acid ceramidase (Ac) deletion has no impact on the course of P. yoelii infection.
(A) The knockout of Ac in T cells was confirmed by analyzing Asah1 mRNA expression of sorted splenic CD4+ and CD8+ T cells from naïve Asah1/Cd4cre/+ (Ac CD4cre knockout [KO]) mice and Asah1/Cd4+/+ littermates (Ac CD4cre wildtype [WT]) as controls via RT-qPCR (n = 2–4). (B) Parasitemia (left panel) and spleen weight (right panel) of P. yoelii-infected Ac CD4cre KO mice and Ac CD4cre WT littermates was determined at indicated time points (n = 7–10). (C) The knockout of Ac in myeloid cells was confirmed by analyzing Asah1 mRNA expression of macrophages, dendritic cells, and neutrophils isolated from spleen, peritoneal lavage (pLavage), and blood of naïve Asah1/Lyz2cre/+ (Ac Lyz2cre KO) mice and Asah1/Lyz2+/+ littermates (Ac Lyz2cre WT) as controls via RT-qPCR (n = 2–6). (D) Parasitemia (left panel) and spleen weight (right panel) of P. yoelii-infected Ac Lyz2cre KO and Ac Lyz2cre WT mice was determined at indicated time points (n = 9). Data from two independent experiments each are presented as mean ± SEM.
-
Figure 3—source data 1
T cell-specific and myeloid-specific acid ceramidase (Ac) deletion has no impact on the course of P. yoelii infection.
- https://cdn.elifesciences.org/articles/77975/elife-77975-fig3-data1-v2.xlsx
Ac KO mice show an impaired erythropoiesis
Since global Ac KO mice showed decreased parasitemia, although cell-specific Ac activity did not alter innate and T cell immune responses during P. yoelii infection per se, we analyzed whether Ac deletion instead has an effect on the host cells of the parasites. Depending on the species, Plasmodium parasites prefer immature or mature RBCs for asexual propagation (Leong et al., 2021). P. yoelii parasites primarily infect reticulocytes, which are defined as Ter119+CD71+ (Martín-Jaular et al., 2013). Therefore, we analyzed frequencies of reticulocytes in blood of Ac WT and Ac KO mice by flow cytometry. Strikingly, reticulocytes were dramatically reduced in blood of noninfected Ac KO mice (Figure 4A, d0). While they were almost absent on days 0 and 3 p.i., frequencies rapidly increased in Ac KO mice 7 days p.i., resulting in equal percentages of reticulocytes in Ac WT and Ac KO mice 10 days p.i. (Figure 4B).
Figure 4 with 1 supplement see all
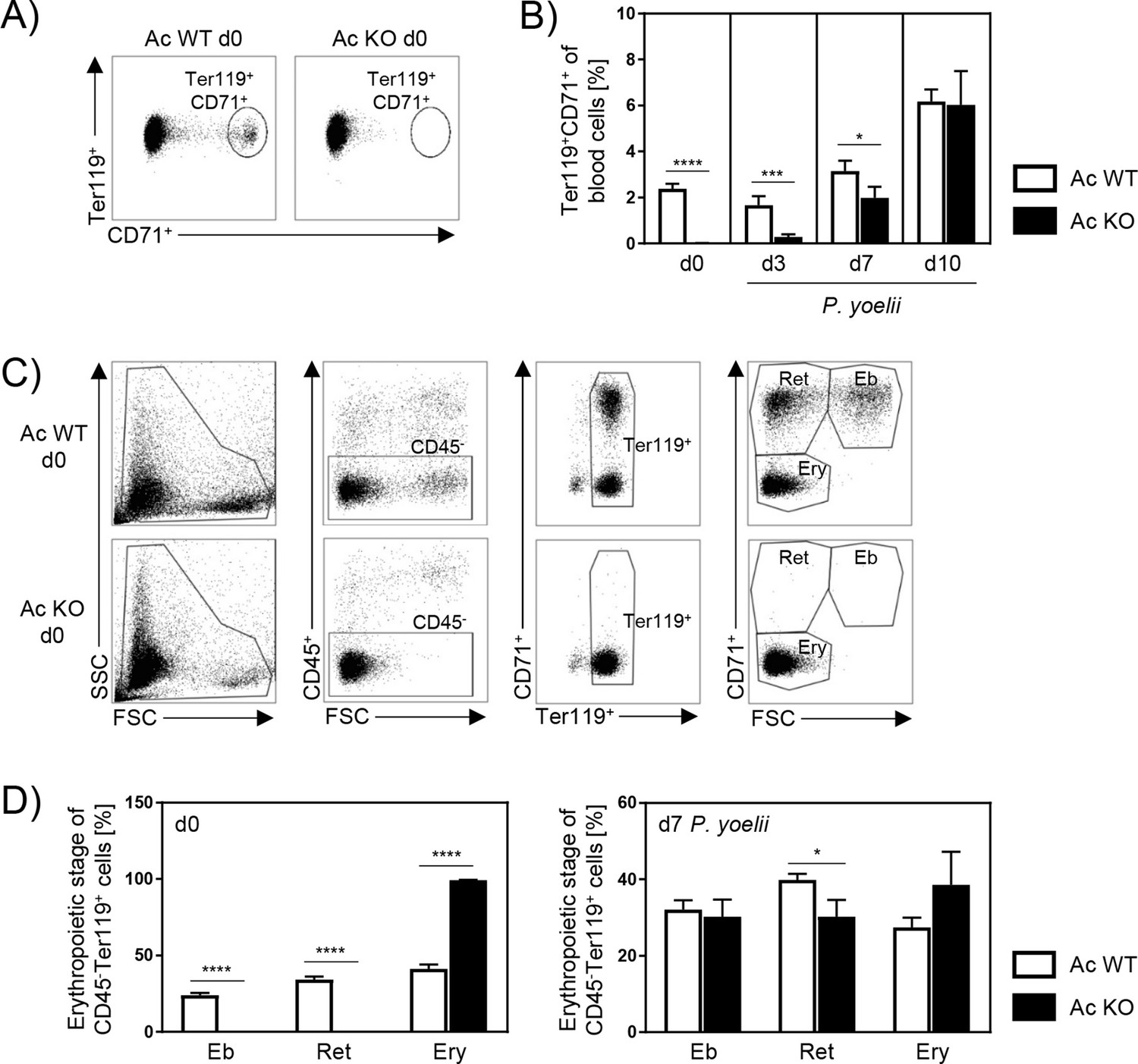
Acid ceramidase (Ac) knockout (KO) affects the erythropoiesis.
(A) Representative flow cytometry dot plot of Ter119+CD71+ reticulocytes in blood of Ac wildtype (WT) and Ac KO mice on day 0. (B) Frequencies of reticulocytes in blood of Ac WT and Ac KO mice were determined by flow cytometry on day 0 and after P. yoelii infection on days 3, 7, and 10 (n = 8–15). (C) Representative flow cytometry gating strategy of erythropoietic developmental stages in bone marrow of Ac WT and Ac KO mice on day 0. Erythropoietic cells were defined as CD45- and Ter119+. Depending on size (forward scatter [FSC]) and CD71 expression, cells were defined as erythroblasts (Eb, CD71+FSChigh), reticulocytes (Ret, CD71+FSClow), and mature erythrocytes (Ery, CD71-FSClow). (D) Frequencies of different erythropoietic stages in bone marrow of noninfected Ac WT and Ac KO mice on day 0 (left panel) and after P. yoelii infection on day 7 (right panel) were analyzed by flow cytometry (n = 6–11). Results from 2–4 independent experiments are presented as mean ± SEM. Statistical analyses were performed using Mann–Whitney U-test for (B) and unpaired Student’s t-test for (D) (*p<0.05, ***p<0.001, ****p<0.0001).
-
Figure 4—source data 1
Acid ceramidase (Ac) knockout (KO) affects the erythropoiesis.
- https://cdn.elifesciences.org/articles/77975/elife-77975-fig4-data1-v2.xlsx
To gain further insights into the impact of Ac expression on erythropoietic cells, we analyzed the development of RBCs within the bone marrow of Ac WT and Ac KO mice in more detail. Depending on size and CD71 expression, erythropoietic cells (CD45-Ter119+) were identified as erythroblasts, reticulocytes, and mature erythrocytes (Figure 4C). As depicted in Figure 4D, erythroblasts and reticulocytes were hardly detectable in bone marrow of noninfected Ac KO mice on day 0 (left panel). Consistent with raising reticulocyte frequencies in the blood (Figure 4B), percentages of erythroblasts and reticulocytes were increasing 7 days p.i. in bone marrow of P. yoelii-infected Ac KO mice, but they were still reduced when compared to infected Ac WT mice (Figure 4D, right panel). Similar results were obtained in noninfected Ac KO mice (Figure 4—figure supplement 1), providing evidence that this phenotype is independent of the infection. Together, these results indicate an impaired erythropoiesis in Ac KO mice, leading to reduced frequencies of reticulocytes the main target cells of P. yoelii parasites.
Ablation of Ac also affects sphingomyelin levels
Our data show that Ac deficiency alters erythropoiesis, resulting in decreased frequencies of reticulocytes in peripheral blood and bone marrow of Ac KO mice. Nonetheless, during P. yoelii infection of Ac KO mice reticulocytes start to repopulate, which is also reflected in rapidly rising parasitemia 10 and 14 days p.i. (Figure 2A). To confirm that the induced Ac KO is persistent at later time points, we analyzed Ac expression in the bone marrow of noninfected mice on days 0 and 7 and 10 days after P. yoelii infection. As validated by RT-qPCR, Ac expression was still absent in Ac KO mice 7 and 10 days p.i. (Figure 5A).
Figure 5 with 1 supplement see all
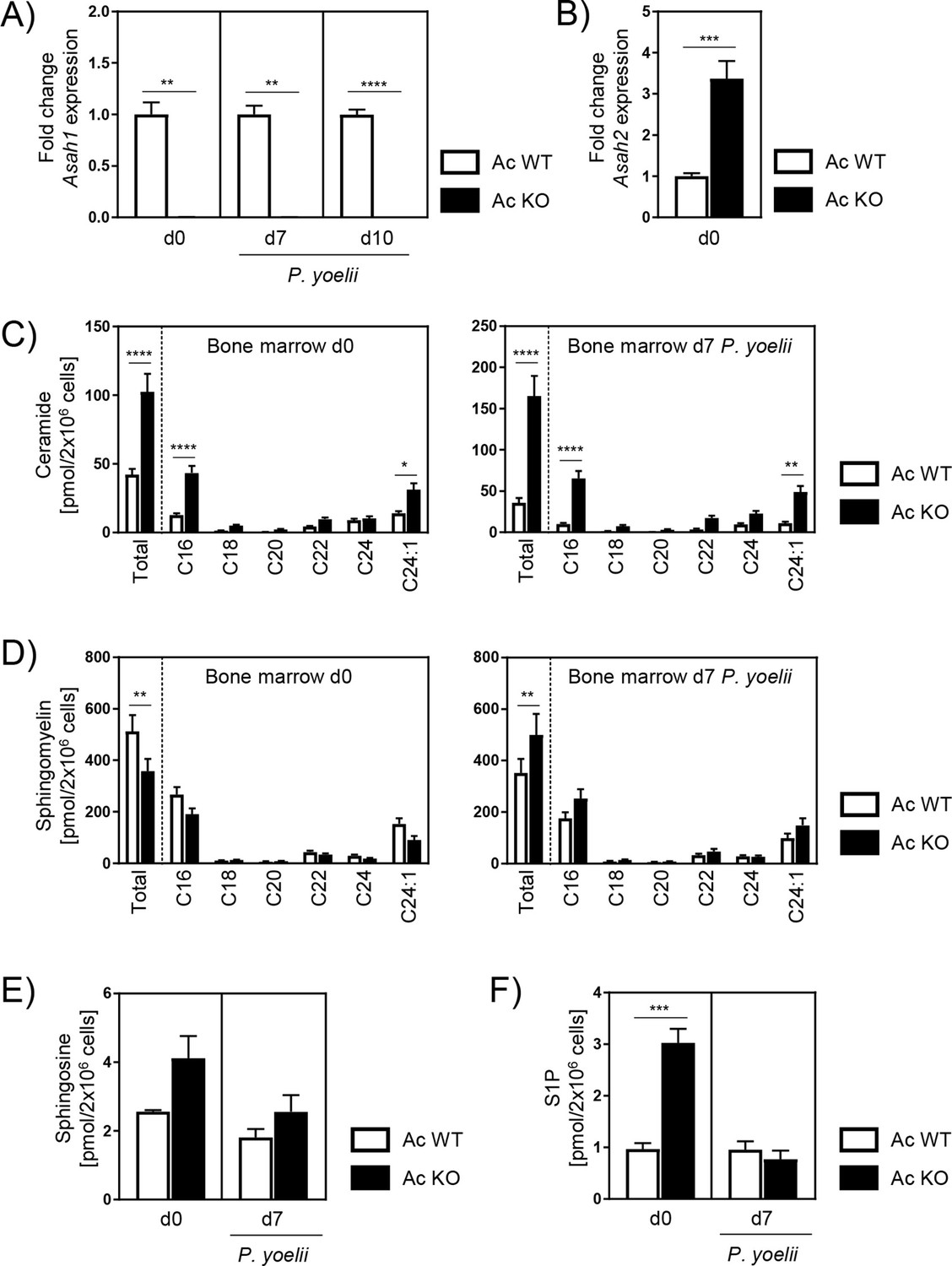
Alterations of sphingolipid metabolism in bone marrow of acid ceramidase (Ac) knockout (KO) mice.
(A) Ac KO validation in bone marrow of noninfected (d0) and P. yoelii-infected (d7 and d10) Ac KO mice and Ac wildtype (WT) littermates as control by RT-qPCR (n = 5–9). (B) Fold change of neutral ceramidase (Asah2) expression in bone marrow on day 0 (n = 6–7). (C) Ceramide, (D) sphingomyelin, (E) sphingosine, and (F) S1P levels in bone marrow of noninfected Ac WT and Ac KO mice on day 0 (left panels), and 7 days post infection (p.i.) (right panels) were determined by high-performance liquid chromatography mass spectrometry (HPLC-MS/MS) (n = 3–6). Data are presented as mean ± SEM. Statistical analyses were performed using Mann–Whitney U-test for (A), unpaired Student’s t-test for (B) and (F), and two-way ANOVA, followed by Sidak’s post-test for (C) and (D) (*p<0.05, **p<0.01, ***p<0.001, ****p<0.0001).
-
Figure 5—source data 1
Alterations of sphingolipid metabolism in bone marrow of acid ceramidase (Ac) knockout (KO) mice.
- https://cdn.elifesciences.org/articles/77975/elife-77975-fig5-data1-v2.xlsx
In order to investigate whether other ceramidases compensate for Ac activity, we analyzed their mRNA expression. We found the neutral ceramidase (Nc) to be upregulated after induction of the Ac KO (Figure 5B). However, analysis of ceramide levels in bone marrow of Ac WT and Ac KO mice revealed significantly higher ceramide concentrations in bone marrow of Ac KO mice compared to Ac WT littermates 7 days p.i. (Figure 5C, right panel), similar to those measured on day 0 (Figure 5C, left panel). This suggests that elevated Nc expression in Ac KO mice is not sufficient to completely compensate for Ac deficiency with regard to the ceramide content.
Next, we determined sphingomyelin, sphingosine, and S1P levels in bone marrow of Ac WT and Ac KO mice. Sphingomyelin is generated from ceramide by the activity of sphingomyelin synthases and has been shown to increase the number of reticulocytes in mice (Scaro et al., 1982). While sphingomyelin levels were decreased in bone marrow of Ac KO mice compared to Ac WT mice on day 0 (Figure 5D, left panel), they were significantly enhanced on day 7 in the presence (Figure 5D, right panel) and absence (Figure 5—figure supplement 1) of the infection, correlating with increasing reticulocyte frequencies observed before. Moreover, Ac KO mice showed slightly increased sphingosine and significantly elevated S1P contents on day 0 in comparison to Ac WT littermates, while levels were similar 7 days p.i. (Figure 5E and F). This might be a consequence of increased expression of neutral ceramidase (Figure 5B). Taken together, these results indicate an altered SL metabolism in bone marrow of Ac KO mice that does not only affect ceramide levels but also leads to significantly elevated sphingomyelin concentrations 7 days p.i., which might account for the repopulation of reticulocytes.
Carmofur treatment leads to alleviated parasitemia and reduced reticulocyte frequencies during P. yoelii infection
Since the genetic ablation of Ac strongly affected erythropoiesis and, as a consequence, the course of P. yoelii infection, we investigated whether pharmacological inhibition of Ac has the same effect. Studies by Realini et al., 2013 showed that the antineoplastic drug carmofur is a potent inhibitor of Ac activity in mice. Therefore, C57BL/6 mice were treated with carmofur or vehicle and infected with P. yoelii (Figure 6A). Strikingly, carmofur-treated mice showed a strong reduction in parasitemia (Figure 6B). Moreover, spleen weights of carmofur-treated mice were significantly reduced on day 7 p.i. (Figure 6C).
Figure 6 with 2 supplements see all
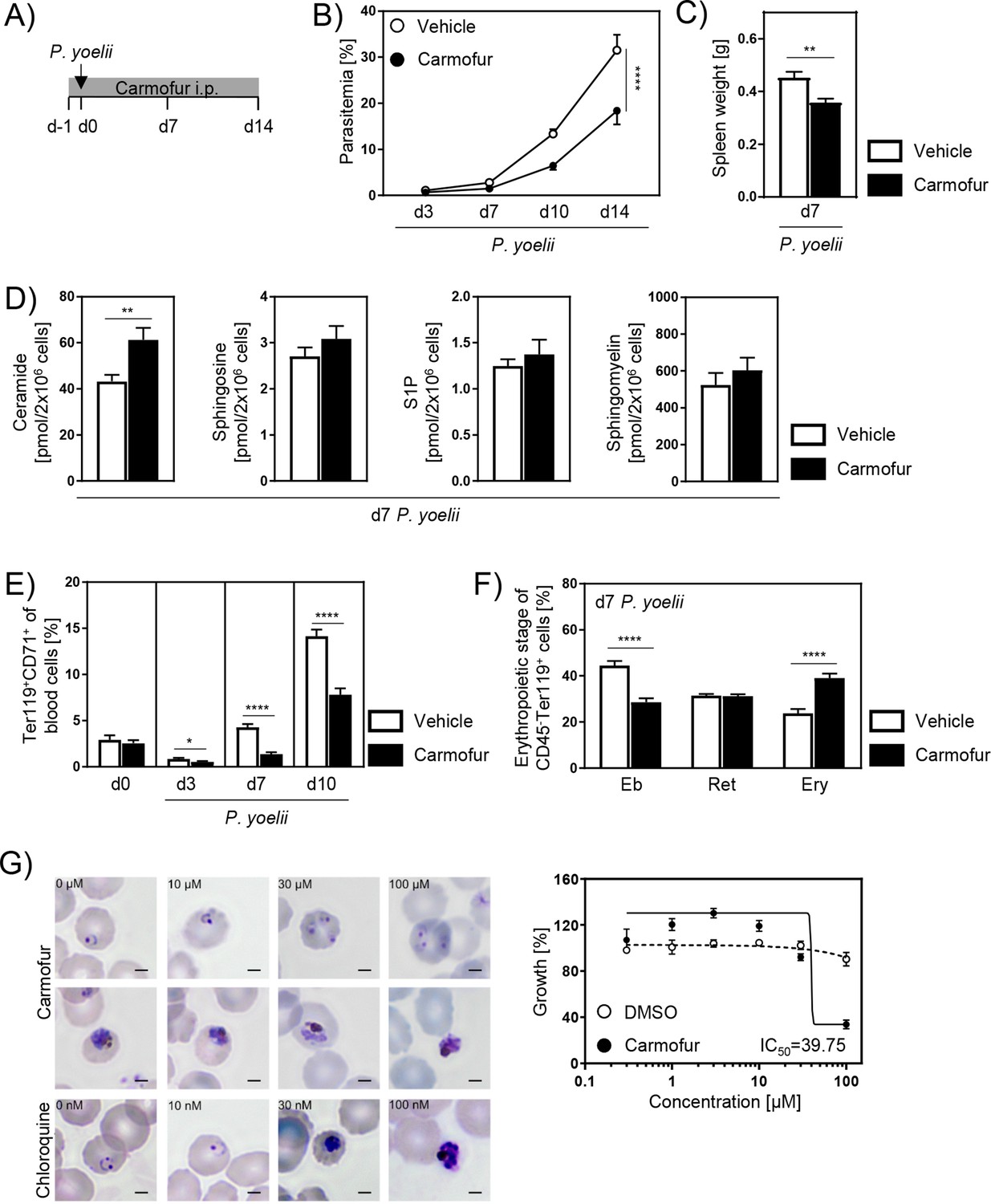
Carmofur treatment leads to decreased parasitemia with lower reticulocyte frequencies.
(A) For pharmacological inhibition of acid ceramidase (Ac), C57BL/6 mice were either treated with 750 µg carmofur or with vehicle as control by daily intraperitoneal (i.p.) injection starting 1 day prior to P. yoelii infection. (B) Parasitemia of infected carmofur- and vehicle-treated mice was determined at indicated time points by microscopy of Giemsa-stained blood films (n = 10–11). (C) Spleen weight of P. yoelii-infected carmofur- and vehicle-treated mice 7 days post infection (p.i.) (n = 9). (D) Ceramide, sphingosine, S1P, and sphingomyelin levels in bone marrow of infected carmofur- and vehicle-treated mice 7 days p.i. were determined by high-performance liquid chromatography mass spectrometry (HPLC-MS/MS) (n = 6–9). (E) Frequencies of Ter119+CD71+ reticulocytes in blood of P. yoelii-infected carmofur- and vehicle-treated mice were analyzed by flow cytometry on days 0, 3, 7, and 10 (n = 12–18). (F) Percentages of erythroblasts (Eb, CD71+FSChigh), reticulocytes (Ret, CD71+FSClow), and mature erythrocytes (Ery, CD71-FSClow) in bone marrow of P. yoelii-infected carmofur- and vehicle-treated mice on day 7 (n = 9). (G) P. falciparum growth inhibition assay to test potential effects of carmofur on intraerythrocytic development in vitro. Micrographs of infected erythrocytes that were either nontreated or exposed 48 hr to increasing doses of carmofur (top rows) or chloroquine (bottom row). Shown are representative images of standard growth assays (left panel). Bars: 2 µm. Dose–response assay (48 hr) of carmofur or DMSO as control against asexual P. falciparum growth (starting at ring stages, n = 3 in triplicates). 50% inhibitory concentration (IC50) is indicated. Results from 2–3 independent experiments are presented as mean ± SEM. Statistical analyses were performed using two-way ANOVA, followed by Sidak’s post-test for (A), unpaired Student’s t-test for (C) and (F), and Mann–Whitney U-test for (D) and (E) (*p<0.05, **p<0.01, ****p<0.0001).
-
Figure 6—source data 1
Carmofur treatment leads to decreased parasitemia with lower reticulocyte frequencies.
- https://cdn.elifesciences.org/articles/77975/elife-77975-fig6-data1-v2.xlsx
In order to confirm Ac inhibition, we determined SL levels in bone marrow of carmofur- and vehicle-treated mice. Indeed, ceramide contents were significantly increased upon carmofur treatment in P. yoelii-infected mice 7 days p.i. (Figure 6D) and noninfected mice (Figure 6—figure supplement 1A) compared to control mice, whereas sphingosine, S1P, and sphingomyelin levels were similar in all groups.
To analyze whether the milder course of infection in carmofur-treated mice is also associated with altered reticulocyte frequencies, we analyzed blood of carmofur- and vehicle-treated mice on day 0 as well as 3, 7, and 10 days after P. yoelii infection. While there is a tendency of decreased reticulocytes on day 0, carmofur-treated mice showed significantly reduced reticulocyte frequencies compared to vehicle-treated mice on days 3, 7, and 10 p.i. (Figure 6E). Furthermore, carmofur treatment led to an altered erythropoiesis in bone marrow with lower relative numbers of erythroblasts and increased percentages of erythrocytes compared to vehicle control 3, 7, and 10 days p.i. (Figure 6F, Figure 6—figure supplement 1D and E). Similar results were obtained in noninfected carmofur- and vehicle-treated mice (Figure 6B–E), indicating that modulation in erythropoiesis occurs independent from the infection.
Plasmodium genomes do not encode candidate acid ceramidases (Aurrecoechea et al., 2009). In support of this notion, cell biological assays and metabolomic profiling revealed ceramide uptake from the infected erythrocyte and modest levels that remain unchanged upon asexual parasite replication (Gulati et al., 2015; Haldar et al., 1991). However, to exclude any potential direct effects of carmofur on intraerythrocytic development in vitro we performed a P. falciparum growth inhibition assay (Figure 6G). Asexual P. falciparum parasites (strain 3D7) were propagated under normal cell culture conditions, enriched for early ring stages and exposed for 48 hr to increasing concentrations of carmofur. As controls, 1% DMSO and the antimalarial drug chloroquine were included. As reported (Vennerstrom et al., 1999), the IC50 concentration of chloroquine in the drug-sensitive 3D7 strain was determined at 22 nM (Figure 6—figure supplement 2). In marked contrast, the IC50 value for carmofur was 39.75 µM, and even at 100 µM viable parasites were detected (Figure 6G, upper panel). For comparison, the IC50 of primaquine, a liver stage-specific drug with no clinical use against blood stages, was reported as 2 µM (Vennerstrom et al., 1999). Moreover, the IC50 of carmofur for Ac was determined at 29 nM (Realini et al., 2013). Hence, we conclude that the effects of carmofur can be solely attributed to the inhibition of host cell Ac.
Carmofur as efficient therapy against P. yoelii infection
Since therapy in clinics is usually started after the infection has already manifested, we next analyzed the therapeutic potential of carmofur treatment. Therefore, we infected C57BL/6 mice with P. yoelii and started with carmofur treatment on day 5 p.i., when the infection was established (Figure 7A). As depicted in Figure 7B, mice showed significantly reduced reticulocyte frequencies after carmofur therapy 7, 10, and 14 days p.i. This was accompanied by a strongly alleviated parasitemia (Figure 7C). Moreover, spleen weights of carmofur-treated mice were decreased compared to vehicle control (Figure 7D). Although carmofur therapy seems to reduce the production of reticulocytes, carmofur-treated mice showed similar or even slightly higher RBC counts than vehicle-treated mice during infection, indicating that they did not suffer from a more severe anemia (Figure 7E).
Figure 7
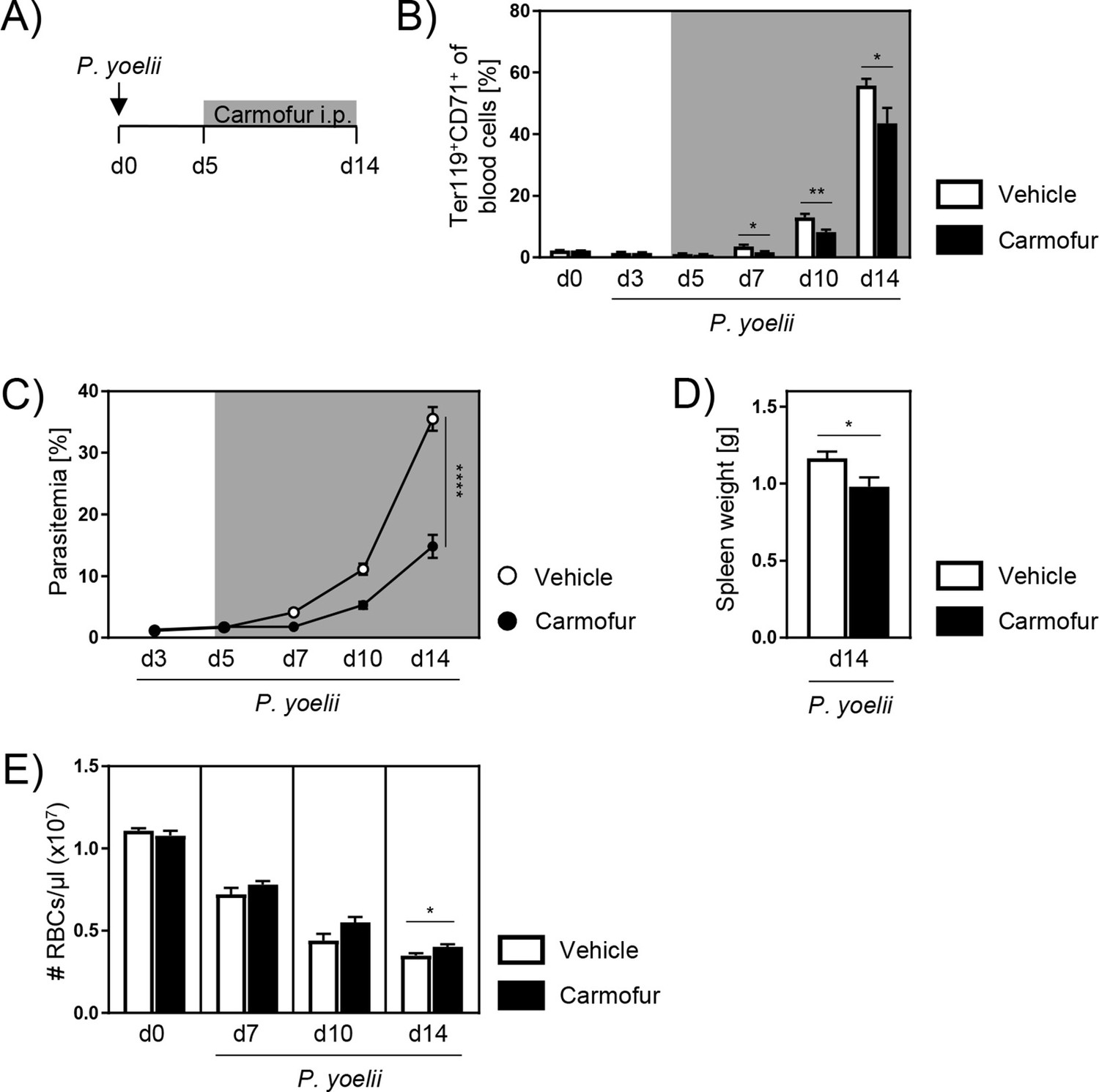
Carmofur application as therapeutic treatment of P. yoelii infection.
(A) As therapeutic approach, C57BL/6 mice were infected with P. yoelii on day 0 and treated with 750 µg carmofur or vehicle only from day 5 post infection (p.i.) onward. (B) Percentages of Ter119+CD71+ reticulocytes in blood were analyzed by flow cytometry at indicated time points (n = 5–10). (C) Parasitemia of infected carmofur- and vehicle-treated mice was determined 3, 5, 7, 10, and 14 days p.i. by microscopy of Giemsa-stained blood films (n = 10). (D) Spleen weight of P. yoelii-infected carmofur- and vehicle-treated mice 14 days p.i. (n = 10). (E) Red blood cell (RBC) count per microliter blood of infected carmofur- and vehicle-treated mice was determined with an automated hematology analyzer (n = 5–10). Results from two independent experiments are summarized as mean ± SEM. Statistical analyses were performed using Mann–Whitney U-test for (B), two-way ANOVA, followed by Sidak’s post-test for (C), and unpaired Student’s t-test for (D) and (E) (*p<0.05, **p<0.01, ****p<0.0001).
-
Figure 7—source data 1
Carmofur application as therapeutic treatment of P. yoelii infection.
- https://cdn.elifesciences.org/articles/77975/elife-77975-fig7-data1-v2.xlsx
In summary, our results provide evidence that deficiency or inhibition of Ac activity alters erythropoiesis, resulting in decreased reticulocyte frequencies associated with lower parasitemia during P. yoelii infection.
Discussion
By hydrolyzing ceramide into sphingosine, Ac is a key regulator of the SL metabolism. We and others showed that ceramide signaling is involved in the regulation of cell differentiation, activation, apoptosis, and proliferation (Hose et al., 2019; Claus et al., 2009; Okazaki et al., 1989). However, the impact of cell-intrinsic Ac activity and ceramide on the course of Plasmodium infection remains unclear. Here, we made use of Ac-deficient mice with ubiquitously increased ceramide levels to study the impact of endogenous ceramide on P. yoelii infection.
An adequate immune response is necessary to fight malaria-causing Plasmodium infections. It is well known that T cells play an important role during the blood stage of malaria (Perez-Mazliah and Langhorne, 2014). Our data show decreased expression of molecules associated with T cell activation in Ac KO mice in the early infection phase (Figure 2D). However, this observation is rather due to a low parasite load than due to reduced T cell activation caused by Ac-deficiency per se. As T cell-specific Ac ablation in Ac CD4cre KO mice did not affect the course of P. yoelii infection (Figure 3B), endogenous ceramide levels do not seem to have a significant impact on T cell activation in this model.
In addition to its characteristics mentioned before, ceramide has also been proposed to modulate erythropoiesis. While long-chain ceramides (C22, C24:0, and C24:1) are described to induce erythroid differentiation in bone marrow cells of rats (Clayton et al., 1974), the treatment with C2 ceramide or bacterial ceramide-generating sphingomyelinase resulted in the inhibition of erythropoiesis in human bone marrow cells (Dallalio et al., 1999). However, these studies were conducted with externally applied ceramide and exclusively in vitro. The role of cell-intrinsic ceramide on RBC development in vivo largely remained elusive. Our data indicate that loss of Ac activity and consequently increased ceramide levels dramatically affect erythrocyte maturation in vivo. Bone marrow of Ac KO mice contained almost no erythroblasts and reticulocytes on day 0 (Figure 4D, left panel). Well in line, studies by Orsini et al., 2019 showed reduced frequencies of orthochromatophilic erythroblasts and absence of reticulocytes in a culture of human hematopoietic stem progenitor cells from umbilical cord blood in response to C2 ceramide or bacterial sphingomyelinase treatment. They propose an inhibition of erythropoiesis independent of apoptosis, mediated by the induction of myelopoiesis via the TNFα/neutral sphingomyelinase/ceramide pathway. In contrast, Signoretto and colleagues observed increased annexin-V staining and therefore elevated eryptosis of human erythrocytes in response to ceramide accumulation induced by treatment with the ceramidase inhibitor ceranib-2 (Signoretto et al., 2016). Although we did not detect a shift from erythropoiesis toward myelopoiesis on day 0 (data not shown), RBCs from Ac KO mice exhibited significantly reduced annexin-V staining compared to Ac WT controls after 24 and 72 hr of in vitro incubation (data not shown). Thus, our data suggest that lower reticulocyte frequencies are not due to increased apoptosis of RBCs. Nevertheless, erythroblasts and reticulocytes seem to repopulate bone marrow and blood of P. yoelii-infected Ac KO mice from day 7 p.i. on (Figure 4B and D, right panel). Since ceramide levels continued to increase until day 7 p.i. (Figure 5C), there was at least no sufficient compensation by other ceramidases, although mRNA expression of Nc was upregulated on day 0 (Figure 5B). However, while sphingomyelin levels were significantly reduced in bone marrow of Ac KO mice compared to Ac WT littermates on day 0 (Figure 5D, left panel), they were highly elevated on day 7 p.i. (Figure 5D, right panel) when erythroblasts and reticulocytes reappear. Sphingomyelin is generated from ceramide by sphingomyelin synthases, which are important regulators of intracellular and plasma membrane ceramide and sphingomyelin levels (Li et al., 2007). Consistent with our data, studies by Clayton et al., 1974 showed an induction of erythropoiesis by sphingomyelin in bone marrow of rats in vitro. Moreover, Scaro and colleagues observed an increase of circulating reticulocytes after injecting mice with purified sphingomyelin in vivo (Scaro et al., 1982). Hence, one might speculate that the massive ceramide accumulation in Ac KO mice leads to an augmented SL turnover, resulting in elevated sphingomyelin concentrations in the bone marrow of Ac KO mice 7 days p.i. that might be responsible for resumption of erythropoiesis.
Several studies provide evidence that SLs and SL analogs are capable of regulating the development of Plasmodium parasites (Labaied et al., 2004; Meyer et al., 2012). C6 ceramide has been shown to efficiently inhibit growth of P. falciparum parasites in a dose-dependent manner in vitro by reducing cytosolic glutathione levels and thereby inducing death of the parasites (Pankova-Kholmyansky et al., 2003). This cytotoxic effect was abolished by addition of S1P, a downstream metabolite of ceramide and sphingosine, respectively. The importance of S1P for parasite survival has recently also been analyzed by Sah et al., 2021. They demonstrated that P. falciparum parasites strictly depend on the erythrocyte endogenous S1P pool for their intracellular development. Inhibition of the S1P-generating enzyme sphingosine kinase-1 resulted in decreased glycolysis, which is essential for parasite energy supply. Interestingly, we measured significantly increased S1P contents in bone marrow of Ac KO mice on day 0 compared to Ac WT littermates, while levels were similar 7 days p.i. (Figure 5F). However, since Ac KO mice show highly reduced parasitemia, we would exclude that enhanced S1P levels beneficially affect parasite growth in our model. It is more likely that the lower parasite load in the early phase of infection is due to highly reduced reticulocyte frequencies in blood of Ac KO mice (Figure 4B), which represent the target cells of P. yoelii parasites (Martín-Jaular et al., 2013). Moreover, after reappearance of the reticulocytes, Ac KO an Ac WT mice exhibited similar parasitemia 10 and 14 days p.i. (Figure 2A).
As our data indicate the involvement of the Ac/ceramide axis in regulating erythropoiesis, which affects the course of Plasmodium infection, we speculated that these findings might open the door to new potential therapeutic strategies against malaria. The limited availability of antimalarial drugs and emerging parasite resistances demand the development of new approaches. Therefore, we made use of carmofur, an antineoplastic drug that has recently been described to inhibit Ac activity in vitro and in vivo (Realini et al., 2013; Dementiev et al., 2019). Carmofur-treated mice exhibited significantly reduced parasitemia and spleen weight compared to control mice during P. yoelii infection (Figure 6B and C). Consistent with our observations in Ac KO mice, carmofur treatment led to decreased reticulocyte frequencies and an altered RBC development (Figure 6E and F). Previous studies have shown the induction of reticulocytosis during the course of P. yoelii infection (Martín-Jaular et al., 2013; Martin-Jaular et al., 2011). As a direct effect of carmofur on the parasites might be possible, the question arises whether the lower parasite load in our model is due to the reduced reticulocyte count or, conversely, whether the lower parasitemia causes the lower reticulocytosis. Using a P. falciparum growth inhibition assay, we could demonstrate that the cytotoxic effect of carmofur is negligible (Figure 6G). Moreover, noninfected carmofur-treated mice show similar changes in erythropoiesis that we observed in the infected animals (Figure 6—figure supplement 1B and C). Hence, we assume that the effects of carmofur are solely attributed to the inhibition of murine Ac. However, possible side effects of carmofur should be considered when thinking about a clinical use as antimalarial therapy. Carmofur has been developed and widely used in Japan to treat patients suffering from carcinomas of the gastrointestinal tract and breast. Like other antineoplastic drugs, it has been reported to increase the risk of leukoencephalopathy, with an incidence of 0.026% between 1982 and 2008 (Mizutani, 2008). Nevertheless, also available antimalarial drugs have adverse effects, including cardiovascular toxicity, myopathy, and neurotoxicity, which should be considered (AlKadi, 2007).
In contrast to the murine model of blood-stage malaria, human Plasmodium infections often go along with low erythropoiesis and reticulocytosis, respectively (Pathak and Ghosh, 2016; Chang et al., 2004a). On the one hand, especially for reticulocyte-restricted species as P. vivax, this leads to a limited number of target cells for the parasites. On the other hand, it results in anemia that causes mortality and morbidity in patients. However, studies by Chang et al. and mathematical calculations by Cromer and colleagues revealed that these beneficial or detrimental effects depend on timing and the rate of reticulocyte production (Chang et al., 2004b; Cromer et al., 2009). Although carmofur-treated mice show a lower reticulocyte production and therefore lower parasitemia, they had similar or even higher RBC counts compared to vehicle-treated controls, meaning that they did not suffer from a more severe anemia (Figure 7E). Thus, carmofur treatment seems to beneficially influence the balance between parasite burden and RBC production. Finally, therapeutic carmofur administration to P. yoelii-infected mice also led to a decreased parasitemia and spleen weight (Figure 7), demonstrating the efficacy of carmofur therapy against malaria, even after manifestation of infection. Nevertheless, it should be considered that carmofur treatment might not be as effective against infections caused by P. falciparum with mixed tropism. Thus, future experiments using rodent Plasmodium chabaudi parasites, which are also able to infect both, reticulocytes and mature erythrocytes (Leong et al., 2021), could clarify this issue.
Overall, our results indicate that cell-intrinsic Ac activity plays an important role during erythropoiesis and therefore during blood-stage infections. Hence, pharmacological inhibition might represent a novel therapeutic strategy to conquer malaria and other infections caused by reticulocyte-prone pathogens.
Materials and methods
Mice
Mice were bred and housed under specific pathogen-free conditions at the animal facility of the University Hospital Essen. All experiments were performed in accordance with the guidelines of the German Animal Protection Law and approved by the State Agency for Nature, Environment, and Consumer Protection (LANUV), North Rhine-Westphalia, Germany (Az 84-02.04.2015.A474, Az 81-02.04.2018.A302). Female C57BL/6 mice were purchased from Envigo Laboratories (Envigo CRS GmbH). Asah1/Rosa26creER mice (Asah1tm1Jhkh/Gt(Rosa)26Sortm9(cre/ESR1)Arte) express a mutated form of the estrogen receptor (ER) that is fused to the Cre recombinase (Taconic Biosciences Inc). Two loxp-sites are flanking exon 1 of the Asah1 gene. Upon tamoxifen administration, the Cre recombinase is translocated into nucleus, leading to the deletion of exon 1 and therefore loss of function. Asah1/Cd4cre mice (Asah1tm1Jhkh/Tg(CD4-cre)1Cwi) were generated by breeding Asah1 mice with Cd4cre mice (Tg(CD4-cre)1Cwi). The Cre recombinase is expressed under the control of CD4 regulatory elements. Asah1/Lyz2cre mice (Asah1tm1Jhkh/Lyz2tm1(cre)Ifo) were obtained by crossing Asah1 mice with Lyz2cre mice (Lyz2tm1(cre)Ifo) that express Cre recombinase under the control of the lysozyme M promoter (Lyz2).
Tamoxifen administration
Request a detailed protocolTamoxifen (Sigma-Aldrich) was dissolved in corn oil and administered to Asah1/Rosa26creER/+ mice (Ac KO) and Asah1/Rosa26+/+ littermates (Ac WT) by intraperitoneal (i.p.) injection (4 mg in 100 µl) 8, 6, and 4 days prior to analysis or infection.
Carmofur treatment
Request a detailed protocolC57BL/6 mice were either treated with 750 µg carmofur (Abcam) dissolved in 100 µl corn oil or with corn oil only (vehicle) by daily i.p. injection starting 1 day prior to infection. For therapeutic use of carmofur, C57BL/6 mice were treated with carmofur or vehicle 5 days p.i.
P. yoelii infection
Request a detailed protocolCryopreserved P. yoelii 17XNL-infected red blood cells (iRBCs) were passaged once through C57BL/6 mice, before using them for infection of experimental mice. Experimental mice were infected with 1 × 105 P. yoelii-parasitized RBCs by intravenous (i.v.) injection on day 0. The parasitemia of infected mice was examined by microscopy of Giemsa-stained blood films. RBC counts were analyzed using the automated hematology analyzer KX-21N (Sysmex).
Cell isolation and flow cytometry
Request a detailed protocolSplenocytes were isolated by rinsing spleens with erythrocyte lysis buffer and washing with PBS supplemented with 2% FCS and 2 mM EDTA. Single-cell suspensions of bone marrow cells were generated by flushing bones (tibia) with PBS. Blood was either taken by puncture of the tail vein during the experiment or by cardiac puncture on day of sacrifice, and immediately mixed with heparin to avoid coagulation. Magnetic-activated cell sorting (MACS) was used to separate and enrich different cell populations from spleen, peritoneal lavage, or blood. Cells of interest were isolated using the CD4+/CD8a+ T cell isolation kit, the Neutrophil isolation kit, CD11c MicroBeads UltraPure, and F4/80 MicroBeads UltraPure (all Miltenyi Biotec) according to the manufacturer’s instructions and separated automatically with the AutoMACS from Miltenyi Biotec. CD4+ and CD8+ T cells were additionally sorted by fluorescence-activated cell sorting (FACS) using an ARIA II cell sorter (BD Bioscience). For flow cytometry analyses, cells were stained with anti-CD4, anti-CD8, anti-CD49d (BD Bioscience), anti-Foxp3, anti-Ki67, anti-CD71 (eBioscience), anti-PD1, anti-CD11a, anti-CD45, and anti-Ter119 (BioLegend) conjugated to fluorescein isothiocyanate (FITC), pacific blue (PB), phycoerythrin (PE), allophycocyanin (APC), Alexa Flour 647 (AF647), or PE-cyanin 7 (PE-Cy7). The fixable viability dye eFluor780 (eBioscience) was used to identify dead cells. Intracellular staining for Foxp3 and Ki67 was performed using the Fixation/Permeabilization kit (eBioscience) according to the manufacturer’s protocol. Cells were analyzed by flow cytometry with an LSR II instrument (BD Bioscience) using DIVA software.
Sphingolipid quantification by HPLC-MS/MS
Request a detailed protocolSpleen tissue homogenates or cell suspensions were subjected to lipid extraction using 1.5 mL methanol/chloroform (2:1, v:v) as described before (Gulbins et al., 2018). The extraction solvent contained d7-sphingosine (d7-Sph), d7-sphingosine 1-phosphate (d7-S1P), C17 ceramide (C17 Cer), and d31-C16 sphingomyelin (d31-C16 SM) (all Avanti Polar Lipids) as internal standards. Chromatographic separations were achieved on a 1290 Infinity II HPLC (Agilent Technologies) equipped with a Poroshell 120 EC-C8 column (3.0 × 150 mm, 2.7 µm; Agilent Technologies). MS/MS analyses were carried out using a 6495 triple-quadrupole mass spectrometer (Agilent Technologies) operating in the positive electrospray ionization mode (ESI+) (Naser et al., 2020). The following mass transitions were recorded (qualifier product ions in parentheses): long-chain bases: m/z 300.3 → 282.3 (252.3) for Sph, m/z 307.3 → 289.3 (259.3) for d7-Sph, m/z 380.3 → 264.3 (82.1) for S1P, and m/z 387.3 → 271.3 (82.1) for d7-S1P; ceramides: m/z 520.5 → 264.3 (282.3) for C16 Cer, m/z 534.5 → 264.3 (282.3) for C17 Cer, m/z 548.5 → 264.3 (282.3) for C18 Cer, m/z 576.6 → 264.3 (282.3) for C20 Cer, m/z 604.6 → 264.3 (282.3) for C22 Cer, m/z 630.6 → 264.3 (282.3) for C24:1 Cer, and m/z 632.6 → 264.3 (282.3) for C24 Cer; sphingomyelins: m/z 703.6 → 184.1 (86.1) for C16 SM, m/z 731.6 → 184.1 (86.1) for C18 SM, m/z 734.6→ 184.1 (86.1) for d31-C16 SM, m/z 759.6 → 184.1 (86.1) for C20 SM, m/z 787.7 → 184.1 (86.1) for C22 SM, m/z 813.7 → 184.1 (86.1) for C24:1 SM, and m/z 815.7 → 184.1 (86.1) for C24 SM. Peak areas of Cer and SM subspecies, as determined with MassHunter software (Agilent Technologies), were normalized to those of the internal standards (C17 Cer or d31-C16 SM) followed by external calibration in the range of 1 fmol to 50 pmol on column. Sph and S1P were directly quantified via their deuterated internal standards d7-Sph (0.25 pmol on column) and d7-S1P (0.125 pmol on column). Determined SL amounts were normalized either to cell count or to the actual protein content (as determined by Bradford assay) of the tissue homogenate used for extraction.
RNA isolation, cDNA synthesis, and qRT-PCR
Request a detailed protocolRNA was either isolated using the RNeasy Fibrous Tissue Kit (QIAGEN) for spleen, thymus, and liver tissue, or using the RNeasy Mini Kit (QIAGEN) for bone marrow cells, T cells, macrophages, dendritic cells, and neutrophils according to the manufacturer’s instructions. To synthesize cDNA, 0.1–1 µg of RNA was reversed transcribed using M-MLV Reverse Transcriptase (Promega) with dNTPs, Oligo-dT mixed with Random Hexamer primers (Thermo Fisher Scientific). For quantitative real-time PCR, Fast SYBR Green Master Mix (Thermo Fisher Scientific) was used on a 7500 Fast Real-Time PCR System (Thermo Fisher Scientific). Each sample was measured as technical duplicate. The expression levels of target genes were normalized against ribosomal protein S9 (RPS9). The following primer sequences (5′–3′) were used: Asah1 (TTC TCA CCT GGG TCC TAG CC, TAT GGT GTG CCA CGG AAC TG), Asah2 (AGA GAG AGC AAG GTA TTC TTC, ACT ATT CAC AAA GTG GTT GC), RPS9 (CTG GAC GAG GGC AAG ATG AAG C, TGA CGT TGG CGG ATG AGC ACA).
P. falciparum growth inhibition assay
Request a detailed protocolGrowth inhibition of cultured P. falciparum was done as published (Smilkstein et al., 2004) with modifications by Spry et al., 2013. Cultures of Pf3D7 master cells were used in flat-bottom 96-well plates. Carmofur (Abcam) was diluted to final concentrations of 0.3, 1, 3, 10, 30, and 100 μM. Chloroquine-diphosphate (Sigma-Aldrich) was diluted to 0.3, 1, 3, 10, 30, and 100 nM final concentrations as positive control. As negative control, the solvent DMSO (1 %) was used. The cultures were adjusted to a parasitemia of 1%, containing predominantly ring stage parasites, assessed by microscopic examination of Giemsa-stained films. The hematocrit was adjusted to the final value of 1% per 200 μl well. Cultures were incubated at 37°C for 48 hr with a gas mixture of 5% O2 and 5% CO2 in N2. After 48 hr, samples were taken for Giemsa-staining and the plates transferred to –80°C for cell lysis. For the fluorescent assay, the plates were thawed and cells were resuspended with 1:1 SYBR Safe DNA Gel Stain (Thermo Fisher, final concentration: 1:10,000) and incubated in the dark at 37°C for 30 min. Fluorescence was measured with a Synergy HTX reader at 485 nm excitation wavelength and 428 nm emission wavelength. To independently confirm the results of the fluorescence assay, parasites were monitored by standard light microscopy of the Giemsa-stained films at 1000× magnification. Arrangement and scaling of the images was done with the Affinity Photo 1.10.5.1342 program.
Statistical analyses
Request a detailed protocolStatistical analyses were performed using GraphPad Prism 7 software. To test for Gaussian distribution, D’Agostino–Pearson omnibus and Shapiro–Wilk normality tests were used. If data passed normality testing unpaired Student’s t-test was performed, otherwise Mann–Whitney U-test was used. Differences between two or more groups with different factors were calculated using two-way ANOVA followed by Sidak’s post-test. p-Values of 0.05 or less were considered indicative of statistical significance (*p<0.05, **p<0.01, ***p<0.001, ****p<0.0001).
Data availability
All data generated or analysed during this study are included in the manuscript and supporting file. Source Data files have been provided for all Figures and Figure Supplements.
References
-
PlasmoDB: a functional genomic database for malaria parasitesNucleic Acids Research 37:D539–D543.https://doi.org/10.1093/nar/gkn814
-
Acid ceramidase rescues cystic fibrosis mice from pulmonary infectionsInfection and Immunity 89:e00677-20.https://doi.org/10.1128/IAI.00677-20
-
Modulation of the course and outcome of blood-stage malaria by erythropoietin-induced reticulocytosisThe Journal of Infectious Diseases 189:735–743.https://doi.org/10.1086/381458
-
Flow cytometric reticulocyte analysis using thiazole orange; clinical experience and technical limitationsClinical and Laboratory Haematology 13:177–188.https://doi.org/10.1111/j.1365-2257.1991.tb00267.x
-
Infection of reticulocytes by Plasmodium falciparum and plasmodium malariae in hyperendemic indigenous malariaAnnals of Tropical Medicine and Parasitology 42:101–112.https://doi.org/10.1080/00034983.1948.11685353
-
Inhibition of sphingomyelin hydrolysis: targeting the lipid mediator ceramide as a key regulator of cellular fateCurrent Medicinal Chemistry 16:1978–2000.https://doi.org/10.2174/092986709788682182
-
Stimulation of erythroblast maturation in vitro by sphingolipidsJournal of Lipid Research 15:557–562.https://doi.org/10.1016/S0022-2275(20)36756-0
-
Low red cell production may protect against severe anemia during a malaria infection--insights from modelingJournal of Theoretical Biology 257:533–542.https://doi.org/10.1016/j.jtbi.2008.12.019
-
Inhibition of human erythroid colony formation by ceramideExperimental Hematology 27:1133–1138.https://doi.org/10.1016/s0301-472x(99)00054-5
-
Molecular mechanism of inhibition of acid ceramidase by carmofurJournal of Medicinal Chemistry 62:987–992.https://doi.org/10.1021/acs.jmedchem.8b01723
-
Human acid ceramidase: processing, glycosylation, and lysosomal targetingThe Journal of Biological Chemistry 276:35352–35360.https://doi.org/10.1074/jbc.M103066200
-
BookMalaria parasites and other haemosporidiaIn: Garnham PC, editors. The American Journal of Tropical Medicine and Hygiene. CABI. pp. 561–563.
-
Use of acid ceramidase and sphingosine kinase inhibitors as antiviral compounds against measles virus infection of lymphocytes in vitroFrontiers in Cell and Developmental Biology 7:218.https://doi.org/10.3389/fcell.2019.00218
-
Antidepressants act by inducing autophagy controlled by sphingomyelin-ceramideMolecular Psychiatry 23:2324–2346.https://doi.org/10.1038/s41380-018-0090-9
-
The accumulation and metabolism of a fluorescent ceramide derivative in Plasmodium falciparum-infected erythrocytesMolecular and Biochemical Parasitology 49:143–156.https://doi.org/10.1016/0166-6851(91)90137-u
-
Principles of bioactive lipid signalling: lessons from sphingolipidsNature Reviews. Molecular Cell Biology 9:139–150.https://doi.org/10.1038/nrm2329
-
Sphingolipids and their metabolism in physiology and diseaseNature Reviews. Molecular Cell Biology 19:175–191.https://doi.org/10.1038/nrm.2017.107
-
Relationship of hepcidin with parasitemia and anemia among patients with uncomplicated Plasmodium falciparum malaria in ghanaThe American Journal of Tropical Medicine and Hygiene 77:623–626.https://doi.org/10.4269/ajtmh.2007.77.623
-
The infection of reticulocytes by plasmodium vivax 1The American Journal of Tropical Medicine and Hygiene s1-18:347–359.https://doi.org/10.4269/ajtmh.1938.s1-18.347
-
Rodent malaria erythrocyte preference assessment by an ex vivo tropism assayFrontiers in Cellular and Infection Microbiology 11:680136.https://doi.org/10.3389/fcimb.2021.680136
-
Inhibition of sphingomyelin synthase (SMS) affects intracellular sphingomyelin accumulation and plasma membrane lipid organizationBiochimica et Biophysica Acta 1771:1186–1194.https://doi.org/10.1016/j.bbalip.2007.05.007
-
Ceramidases: regulators of cellular responses mediated by ceramide, sphingosine, and sphingosine-1-phosphateBiochimica et Biophysica Acta 1781:424–434.https://doi.org/10.1016/j.bbalip.2008.06.002
-
Sphingolipid analogues inhibit development of malaria parasitesACS Medicinal Chemistry Letters 3:43–47.https://doi.org/10.1021/ml2002136
-
Leukoencephalopathy caused by antineoplastic drugsBrain and Nerve = Shinkei Kenkyu No Shinpo 60:137–141.
-
From erythroblasts to mature red blood cells: organelle clearance in mammalsFrontiers in Physiology 8:1076.https://doi.org/10.3389/fphys.2017.01076
-
Metabolic regulation of hematopoietic stem cell commitment and erythroid differentiationCurrent Opinion in Hematology 23:198–205.https://doi.org/10.1097/MOH.0000000000000234
-
Sphingomyelin turnover induced by vitamin D3 in HL-60 cellsJournal of Biological Chemistry 264:19076–19080.https://doi.org/10.1016/S0021-9258(19)47268-2
-
Ceramide mediates growth inhibition of the Plasmodium falciparum parasiteCellular and Molecular Life Sciences 60:577–587.https://doi.org/10.1007/s000180300049
-
CD4 T-cell subsets in malaria: TH1/TH2 revisitedFrontiers in Immunology 5:671.https://doi.org/10.3389/fimmu.2014.00671
-
Host-mediated regulation of superinfection in malariaNature Medicine 17:732–737.https://doi.org/10.1038/nm.2368
-
Ceranib-2-induced suicidal erythrocyte deathCell Biochemistry and Function 34:359–366.https://doi.org/10.1002/cbf.3196
-
Simple and inexpensive fluorescence-based technique for high-throughput antimalarial drug screeningAntimicrobial Agents and Chemotherapy 48:1803–1806.https://doi.org/10.1128/AAC.48.5.1803-1806.2004
-
Iron, anemia and hepcidin in malariaFrontiers in Pharmacology 5:125.https://doi.org/10.3389/fphar.2014.00125
-
8-aminoquinolines active against blood stage Plasmodium falciparum in vitro inhibit hematin polymerizationAntimicrobial Agents and Chemotherapy 43:598–602.https://doi.org/10.1128/AAC.43.3.598
-
Hepcidin is regulated during blood-stage malaria and plays a protective role in malaria infectionJournal of Immunology 187:6410–6416.https://doi.org/10.4049/jimmunol.1101436
-
Acid ceramidase deficiency: farber disease and SMA-PMEOrphanet Journal of Rare Diseases 13:121.https://doi.org/10.1186/s13023-018-0845-z
Article and author information
Author details
Astrid M Westendorf

Funding
Deutsche Forschungsgemeinschaft (GRK2098)
- Karl Sebastian Lang
Deutsche Forschungsgemeinschaft (GRK2581)
- Burkhard Kleuser
The funders had no role in study design, data collection and interpretation, or the decision to submit the work for publication.
Acknowledgements
We kindly thank Sina Luppus and Christina Liebig for excellent technical assistance. We further thank Daniel Herrmann for the help with HPLC-MS/MS sphingolipid quantification. This work was supported by the Deutsche Forschungsgemeinschaft (DFG-GRK2098 to AMW, JB, KSL, EG, WH, and DFG-GRK2581 to BK).
Ethics
All experiments were performed in strict accordance with the guidelines of the German Animal Protection Law and approved by the State Agency for Nature, Environment, and Consumer Protection (LANUV), North Rhine-Westphalia, Germany (Az 84-02.04.2015.A474, Az 81-02.04.2018.A302).
Copyright
© 2022, Günther et al.
This article is distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use and redistribution provided that the original author and source are credited.
Metrics
-
- 796
- views
-
- 222
- downloads
-
- 2
- citations
Views, downloads and citations are aggregated across all versions of this paper published by eLife.
Download links
A two-part list of links to download the article, or parts of the article, in various formats.
Downloads (link to download the article as PDF)
Open citations (links to open the citations from this article in various online reference manager services)
Cite this article (links to download the citations from this article in formats compatible with various reference manager tools)
The acid ceramidase/ceramide axis controls parasitemia in Plasmodium yoelii-infected mice by regulating erythropoiesis
eLife 11:e77975.
https://doi.org/10.7554/eLife.77975
Further reading
-
- Microbiology and Infectious Disease
The persistence of latent viral reservoirs remains the major obstacle to eradicating human immunodeficiency virus (HIV). We herein found that ICP34.5 can act as an antagonistic factor for the reactivation of HIV latency by herpes simplex virus type I (HSV-1), and thus recombinant HSV-1 with ICP34.5 deletion could more effectively reactivate HIV latency than its wild-type counterpart. Mechanistically, HSV-ΔICP34.5 promoted the phosphorylation of HSF1 by decreasing the recruitment of protein phosphatase 1 (PP1α), thus effectively binding to the HIV LTR to reactivate the latent reservoirs. In addition, HSV-ΔICP34.5 enhanced the phosphorylation of IKKα/β through the degradation of IκBα, leading to p65 accumulation in the nucleus to elicit NF-κB pathway-dependent reactivation of HIV latency. Then, we constructed the recombinant HSV-ΔICP34.5 expressing simian immunodeficiency virus (SIV) env, gag, or the fusion antigen sPD1-SIVgag as a therapeutic vaccine, aiming to achieve a functional cure by simultaneously reactivating viral latency and eliciting antigen-specific immune responses. Results showed that these constructs effectively elicited SIV-specific immune responses, reactivated SIV latency, and delayed viral rebound after the interruption of antiretroviral therapy (ART) in chronically SIV-infected rhesus macaques. Collectively, these findings provide insights into the rational design of HSV-vectored therapeutic strategies for pursuing an HIV functional cure.
-
- Biochemistry and Chemical Biology
- Microbiology and Infectious Disease
Teichoic acids (TA) are linear phospho-saccharidic polymers and important constituents of the cell envelope of Gram-positive bacteria, either bound to the peptidoglycan as wall teichoic acids (WTA) or to the membrane as lipoteichoic acids (LTA). The composition of TA varies greatly but the presence of both WTA and LTA is highly conserved, hinting at an underlying fundamental function that is distinct from their specific roles in diverse organisms. We report the observation of a periplasmic space in Streptococcus pneumoniae by cryo-electron microscopy of vitreous sections. The thickness and appearance of this region change upon deletion of genes involved in the attachment of TA, supporting their role in the maintenance of a periplasmic space in Gram-positive bacteria as a possible universal function. Consequences of these mutations were further examined by super-resolved microscopy, following metabolic labeling and fluorophore coupling by click chemistry. This novel labeling method also enabled in-gel analysis of cell fractions. With this approach, we were able to titrate the actual amount of TA per cell and to determine the ratio of WTA to LTA. In addition, we followed the change of TA length during growth phases, and discovered that a mutant devoid of LTA accumulates the membrane-bound polymerized TA precursor.




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}