Evidence for a role of human blood-borne factors in mediating age-associated changes in molecular circadian rhythms
- Howard Hughes Medical Institute, Perelman School of Medicine, University of Pennsylvania, United States
- Chronobiology and Sleep Institute, Perelman School of Medicine, University of Pennsylvania, United States
- Institute for Translational Medicine and Therapeutics (ITMAT), Perelman School of Medicine, University of Pennsylvania, United States
- Department of Genetics, Perelman School of Medicine, University of Pennsylvania, United States
Figures
Figure 1 with 2 supplements
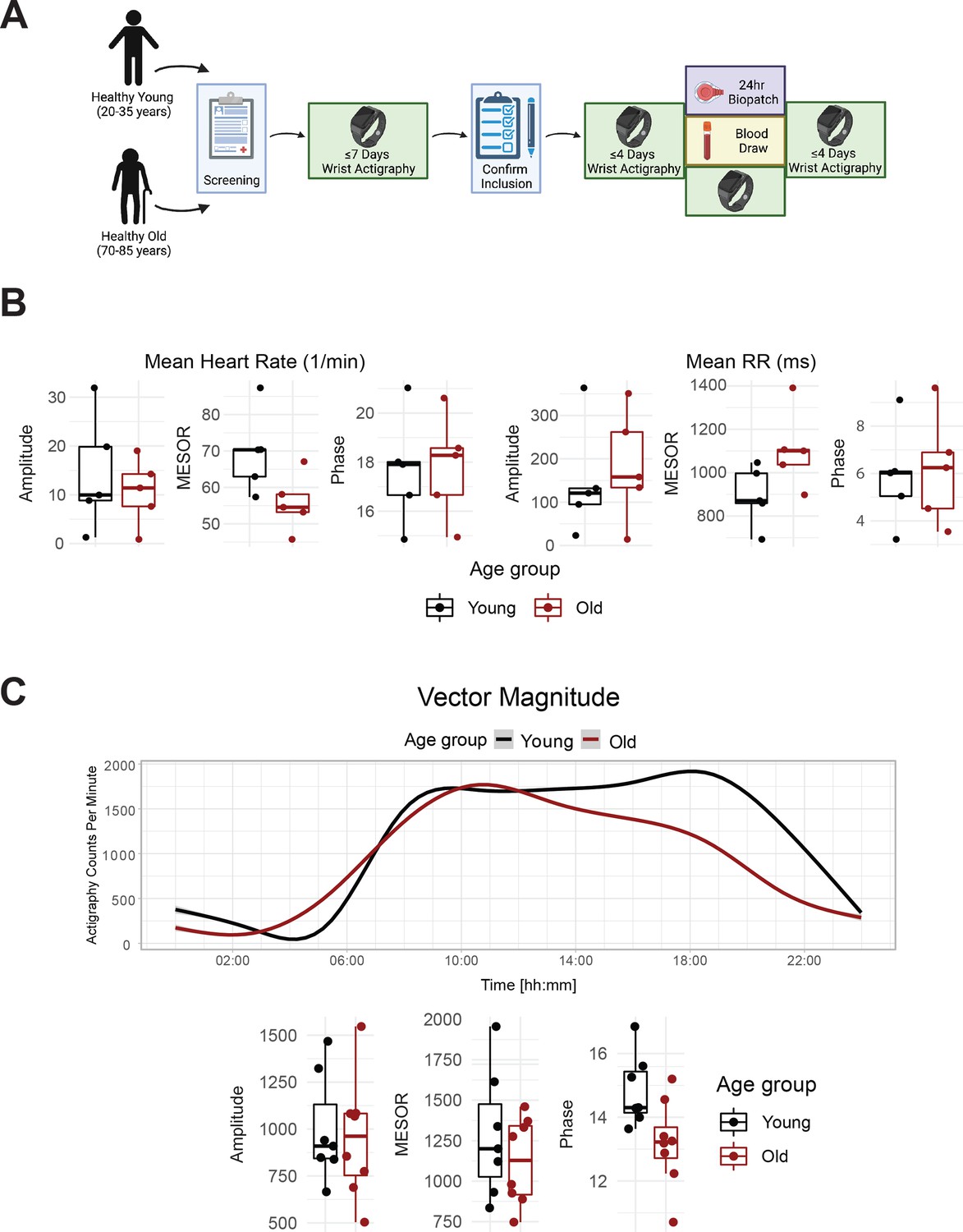
Healthy elderly individuals tend to have lower heart rate, increased heart rate variability, and phase advanced activity patterns relative to young subjects.
Experimental protocol for enrollment and monitoring of human subjects (A). As assessed by Zyphyr BioPatch, electrocardiogram (EKG) measurements suggest that MESOR of average heart rate decreases with age (p=0.056) and that MESOR of heart rate variability increases with age (p=0.056). N=5 per group. MESOR and amplitude were tested using two-sided Wilcoxon rank sum exact test, while phase was tested by Kuiper’s two-sample test (B). Average activity counts across three axes (vector magnitude), as recorded by the actigraph device plotted throughout the day (top) and analyzed for circadian rhythm (bottom) (C). While the amplitudes of activity did not differ between the age groups, the older individuals trended towards an early phase (p~0.055, Kupier’s two-sample test) compared to young individuals. N=7 for young and N=8 for old. Lines in the top panel of C are smoothed means (fit with penalized cubic regression splines) for data from each age group. Dots in the bottom panel of C are subject-level cosinor parameter estimates derived from cosinor fits to the actigaraphy data. Boxplot midlines correspond to median values, while the lower and upper hinges correspond to the first and their quartiles, respectively. Boxplot whiskers extend to the smallest/largest points within 1.5 * IQR (Inter Quartile Range) of the lower/upper hinge. Panel A was created with BioRender.com.
-
Figure 1—source data 1
A flow chart of the number of subjects included in each analysis present in this study.
- https://cdn.elifesciences.org/articles/88322/elife-88322-fig1-data1-v1.xlsx
Figure 1—figure supplement 1
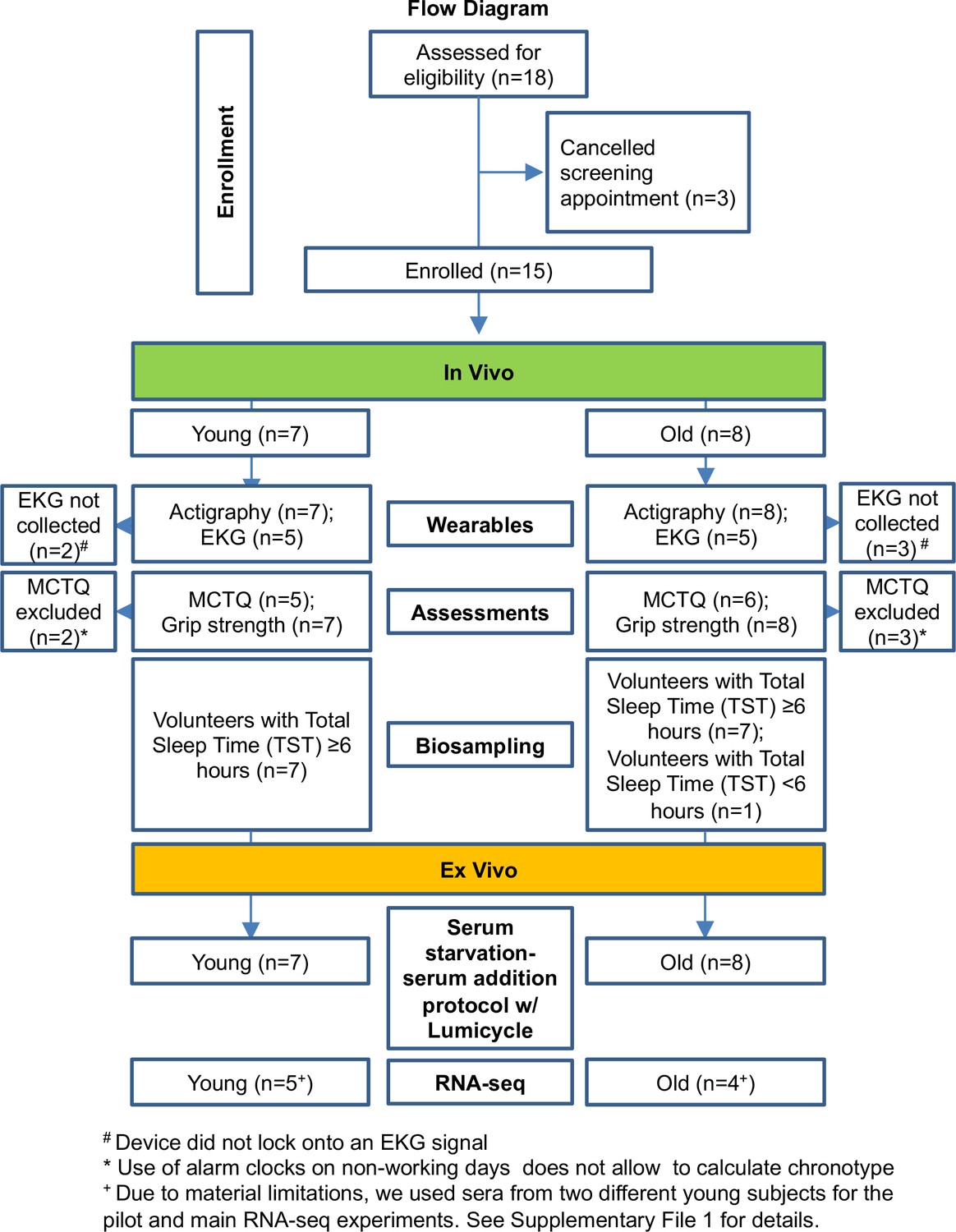
Subject inclusion flow chart.
A flow chart of the number of subjects included in each analysis present in this study.
Figure 1—figure supplement 2
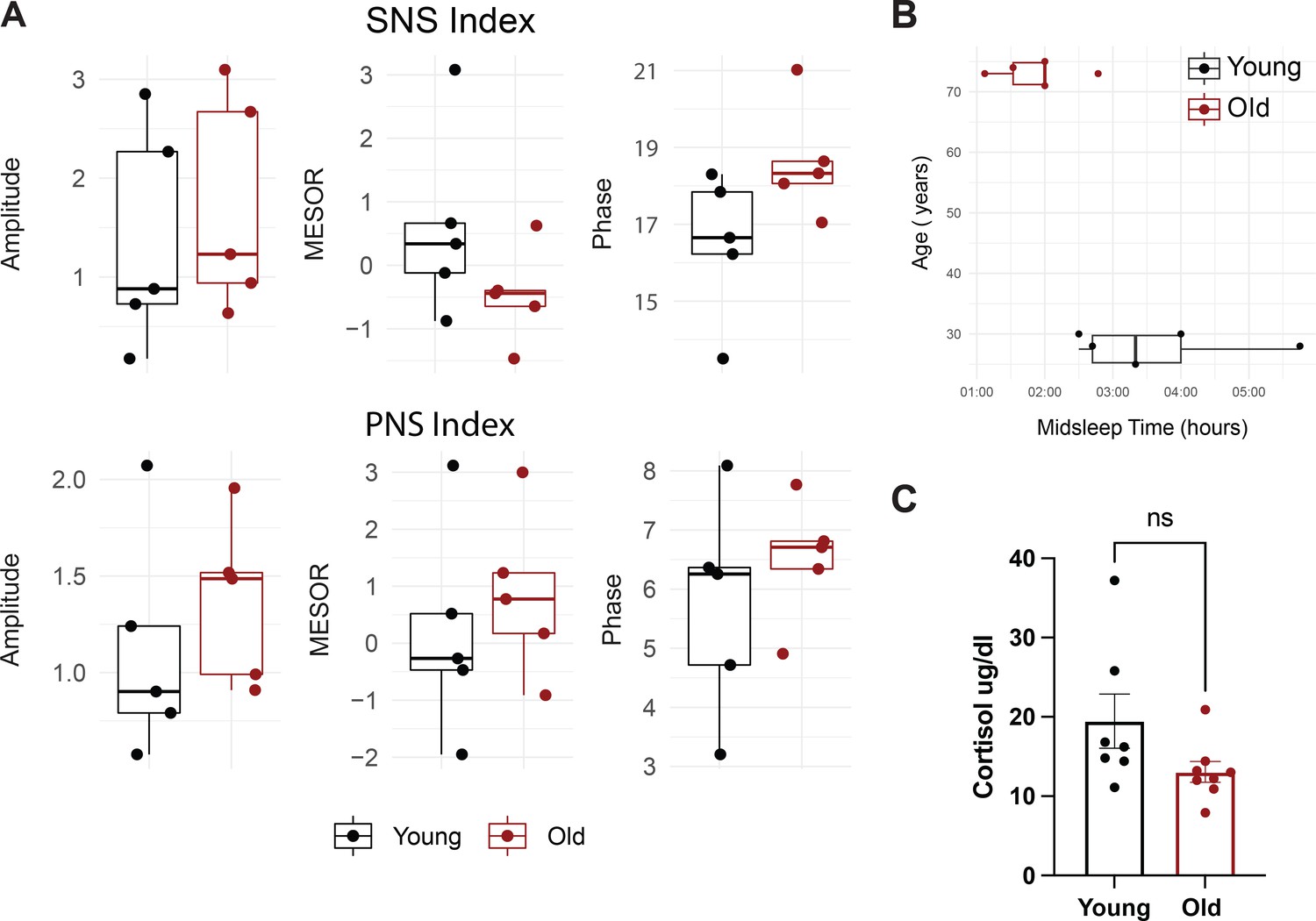
There are differences in midsleep time, but no significant differences in sympathetic, parapsympathetic nervous system indices, or cortisol levels between young and old individuals.
Sympathetic nervous system (SNS, A, top) and parasympathetic nervous system (PNS, A, bottom) indices were measured using EKG. Amplitude, MESOR, and Phase were calculated from the resulting oscillations and the Median, quartiles, and SEMs are shown. Dots represent individuals. p>0.05. N=5 per group. MESOR and amplitude were tested using Wilcoxon rank sum exact test, while phase was tested by Kuiper’s two-sample test. Boxplot midlines correspond to median values, while the lower and upper hinges correspond to the first and third quartiles, respectively. Boxplot whiskers extend to the smallest/largest points within 1.5 * IQR (inter Quartile Range) of the lower/upper hinge (A). Boxplot distributions of midsleep times for subjects in the young (black) and old (red) cohorts. The midsleep time is advanced in the old subjects compared to young by Mann-Whitney U test (Wilcoxon rank sum test p=0.036). The upper whisker extends from the hinge to the largest value no further than 1.5 * IQR from the hinge (where IQR is the inter-quartile range, or distance between the first and third quartiles). The lower whisker extends from the hinge to the smallest value at most 1.5 * IQR of the hinge. We calculated midsleep times for each subject from their responses to the Munich Chronotype Questionnaire (MCTQ). N=5 per group. We were not able to calculate midsleep times for four subjects (two from the old cohort and two from the young cohort) because they used alarms to wake on their non-working days and one old subject because they exhibited a total sleep time less than 6 hr (B). Serum cortisol levels are not statistically different by unpaired t-test (C) Summary statistics are displayed as mean +/- SEM. (N=7 young, 8 old).4.
-
Figure 1—figure supplement 2—source data 1
Raw data obtained from serum cortisol measurements.
- https://cdn.elifesciences.org/articles/88322/elife-88322-fig1-figsupp2-data1-v1.xls
Figure 2 with 4 supplements
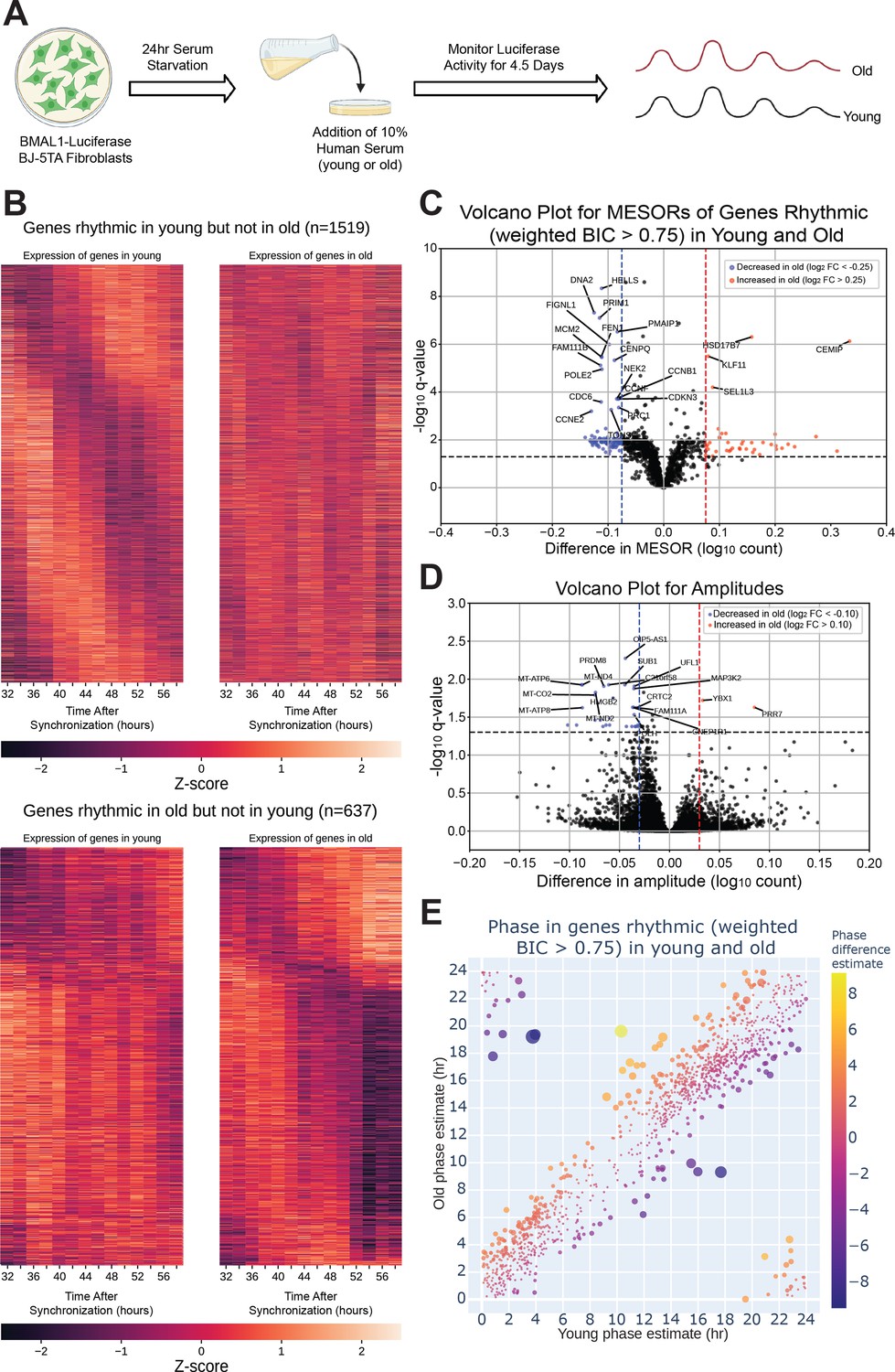
The circadian transcriptome is differentially affected by entrainment with sera from young and old human subjects.
A visual representation of the serum starvation-serum addition protocol to synchronize the BMAL1-luciferase BJ-5TA fibroblasts (A). When comparing the circadian transcriptome entrained by either young or old sera (n=4 sera per group), 1519 genes lost rhythmicity with age (B, top) while only 637 genes gained rhythmicity with age (B, bottom). Weighted BIC criterion with a threshold of 0.75 was used to assess rhythmicity (B). The number of genes rhythmic in young and old (according to weighted BIC > 0.75 criterion) that show a detectable change in MESOR (q<0.05 criterion for MESOR difference) is 568, with MESOR increasing in 163 and decreasing in 405 genes. Out of these, 40 genes with increased MESOR (red) and 98 genes with decreased MESOR (blue) also satisfy the condition |log₂ FC|> 0.25 (C). We were only able to detect change in amplitude for a small number of genes, 39 genes had decreased amplitude in old and 2 had increased amplitude using CircaCompare. Only 30 genes with decreased amplitude also satisfy the condition |log₂ FC|> 0.1 (blue) (D). For phase, using a test provided by CircaCompare, under q<0.05 cutoff for age-related phase differences in genes rhythmic in young and old (with BIC > 0.75 cycling criteria), we detected 20 genes with advanced phase, and 34 with delayed phase (E). Panel A was created with BioRender.com.
Figure 2—figure supplement 1
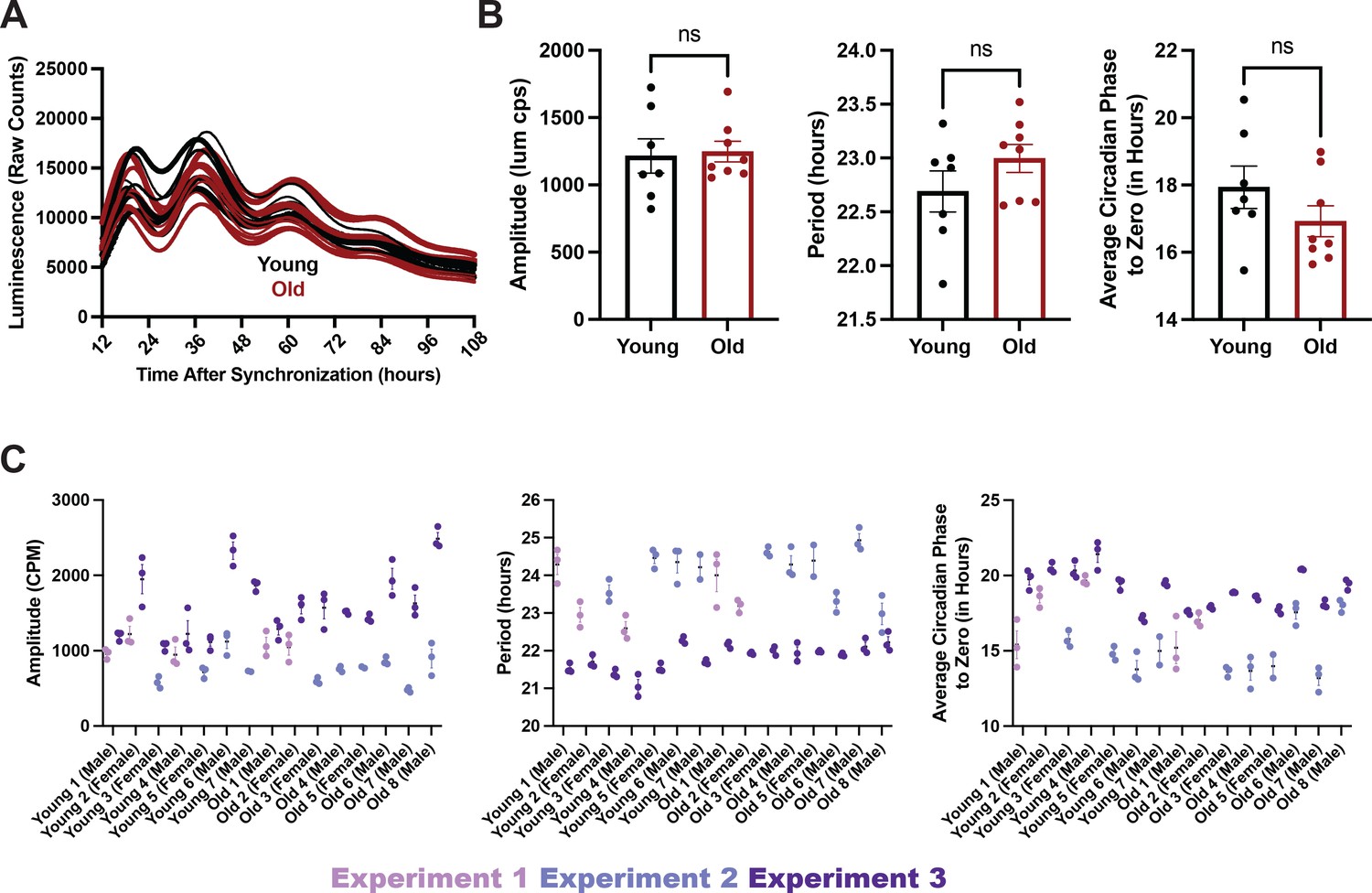
Young and old serum are equally effective at entraining cells in culture.
(A, B) Cells synchronized with human serum from either young (N=7 subjects) or old (N=8 subjects) individuals did not show differences in amplitude (as measured in luminescence counts per second, bottom, left), period (bottom, middle), or phase (bottom, right). Line traces and data points represent average values for a specific patient’s serum over 2 experiments with 2–3 replicates per experiment. A visual representation of the individual replicates averaged in B (C) Summary statistics are displayed as mean +/- SEM. Means compared by unpaired t-test. Despite individual and run to run variability, cells synchronized with human serum from either young (N=7 subjects) or old (N=8 subjects) individuals showed similar amplitude (left), period (middle), and phase relative to synchronization time (right) of BMAL1-luciferase rhythms. Data points represent individual replicates within an experiment. Wells run in the same experiment are displayed in the same color. Each subject’s sample was run in 2–3 replicates.
-
Figure 2—figure supplement 1—source data 1
Circadian transcripome analysis of Day 2 of serum entrainment compared to Day 1.
- https://cdn.elifesciences.org/articles/88322/elife-88322-fig2-figsupp1-data1-v1.xlsx
Figure 2—figure supplement 2
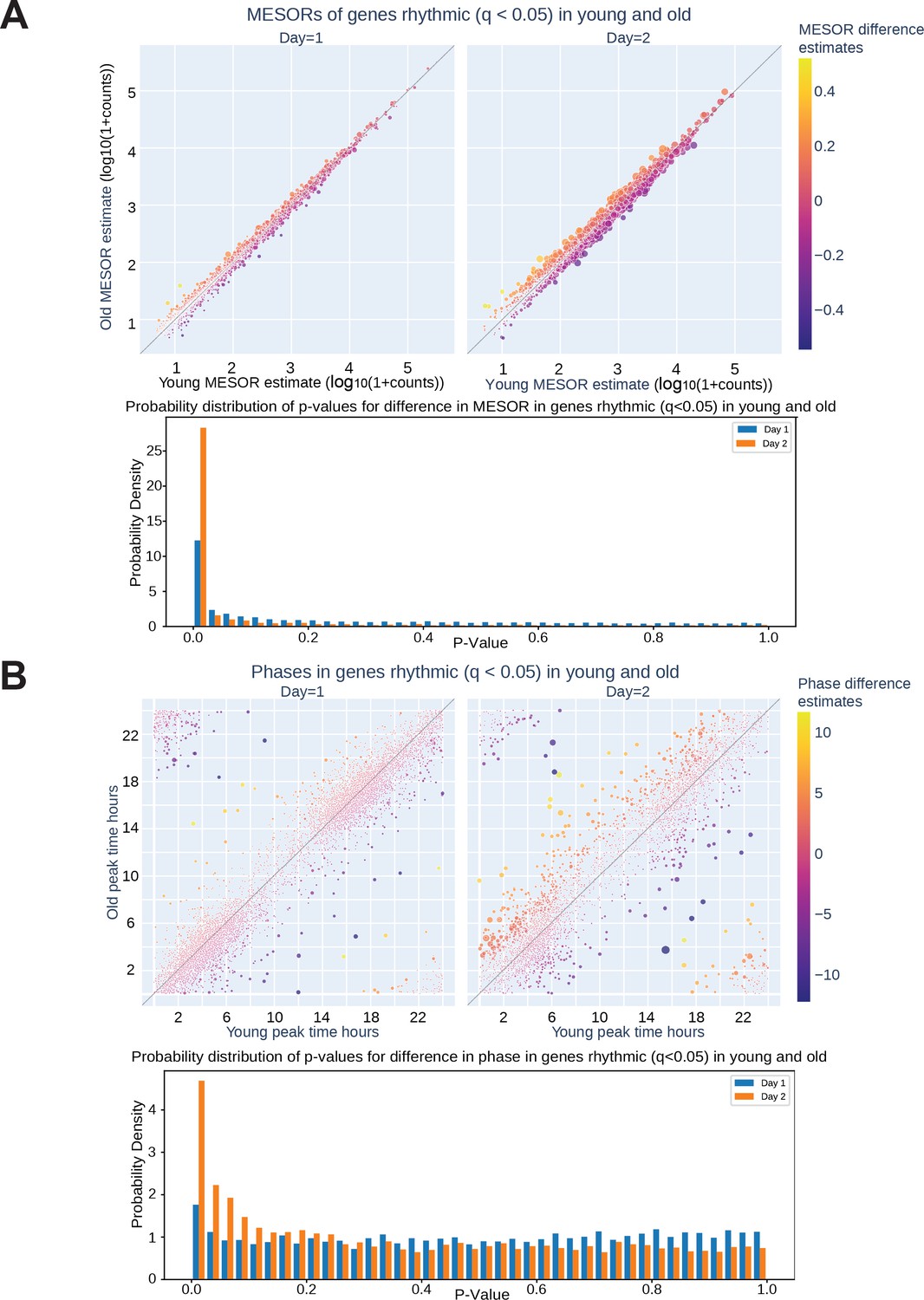
The circadian transcriptomes of cells synchronized with young or old sera deviate significantly on Day 2.
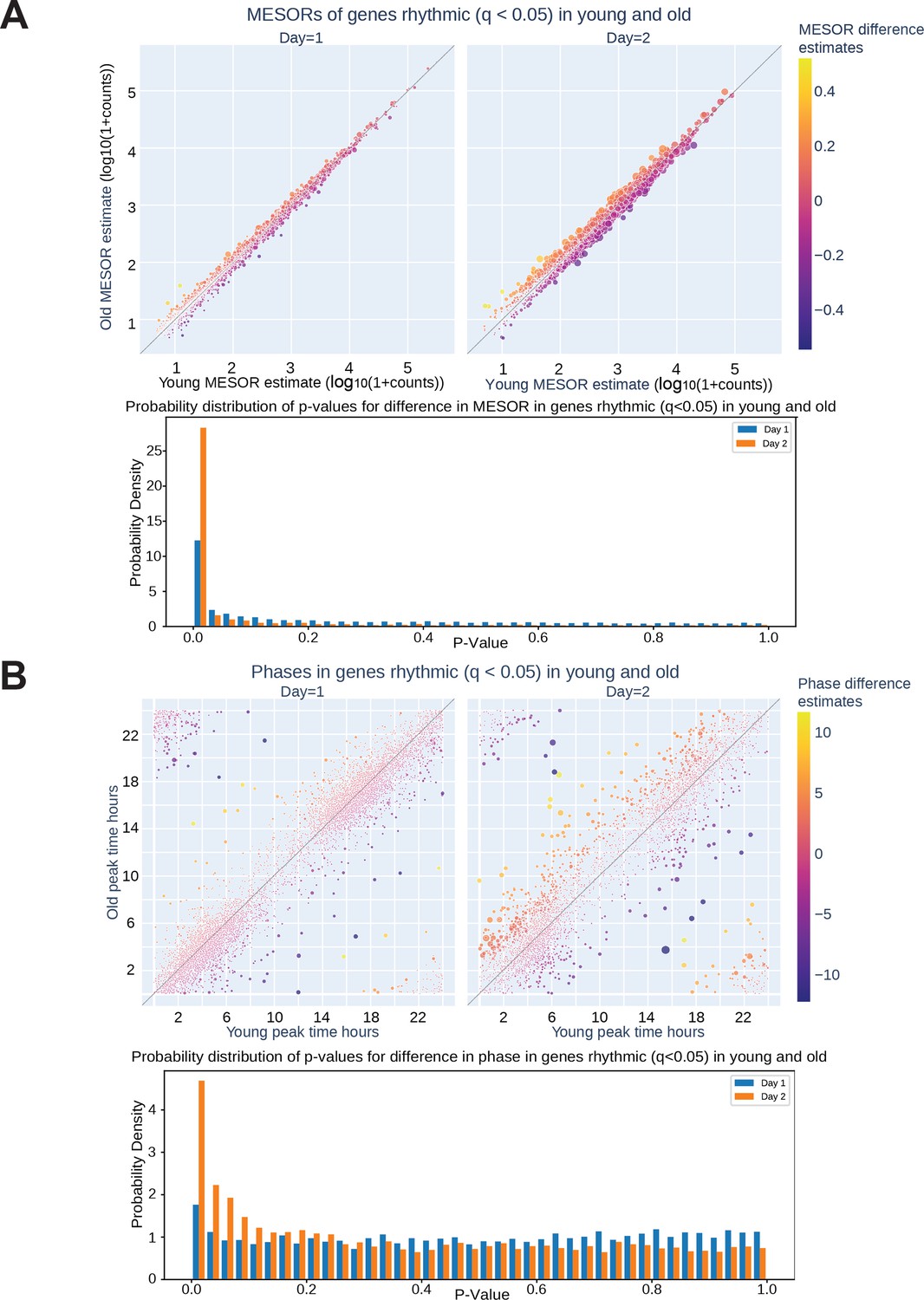
On Day 2 of serum entrainment, young and old transcriptional rhythms differentiated. CircaCompare analysis of RNA sequencing revealed that on Day 2 of serum entrainment analysis (36–58 hr after synchronization) the MESOR differences of cycling genes are larger between the cells entrained with young or old serum (A, top). Additionally, more genes were phase shifted in the old serum condition compared to young serum on Day 2 (B, top). The distribution of p-values shows an enrichment of low p-values on Day 2 for both MESOR and phase differences (A, bottom, B, bottom). The size of each circle in the scatter plots is proportional to -log10q, hence bigger circles correspond to smaller q-values for the difference between young and old for each metric.
Figure 2—figure supplement 3
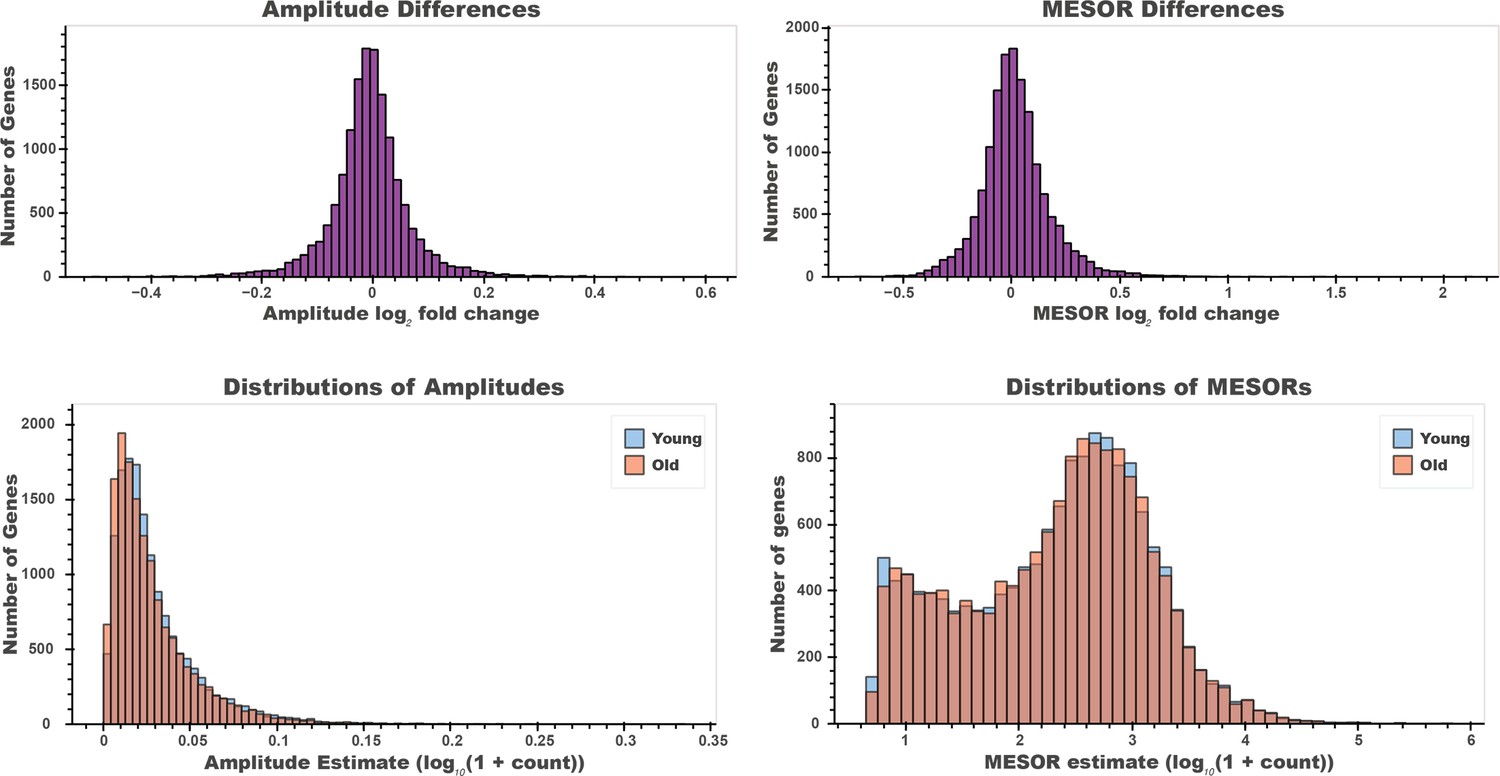
There is a small effect of age on the amplitude and MESOR of cycling mRNA between young and old serum samples.
The difference (Top, left) between young and old mRNA amplitude values (bottom, left) are significantly different by Wilcoxon signed-rank test (p=3.73e-39). The difference (Top, right) between young and old mRNA MESOR values (bottom, right) are significantly different by Wilcoxon signed-rank test (p=6.94e-22). However, these small differences result in very low p-values due to the large number of genes inspected.
Figure 2—figure supplement 4
Sample traces of transcripts that cycle with young serum synchronization and are differentially affected by old serum synchronization.
Figure 3 with 1 supplement
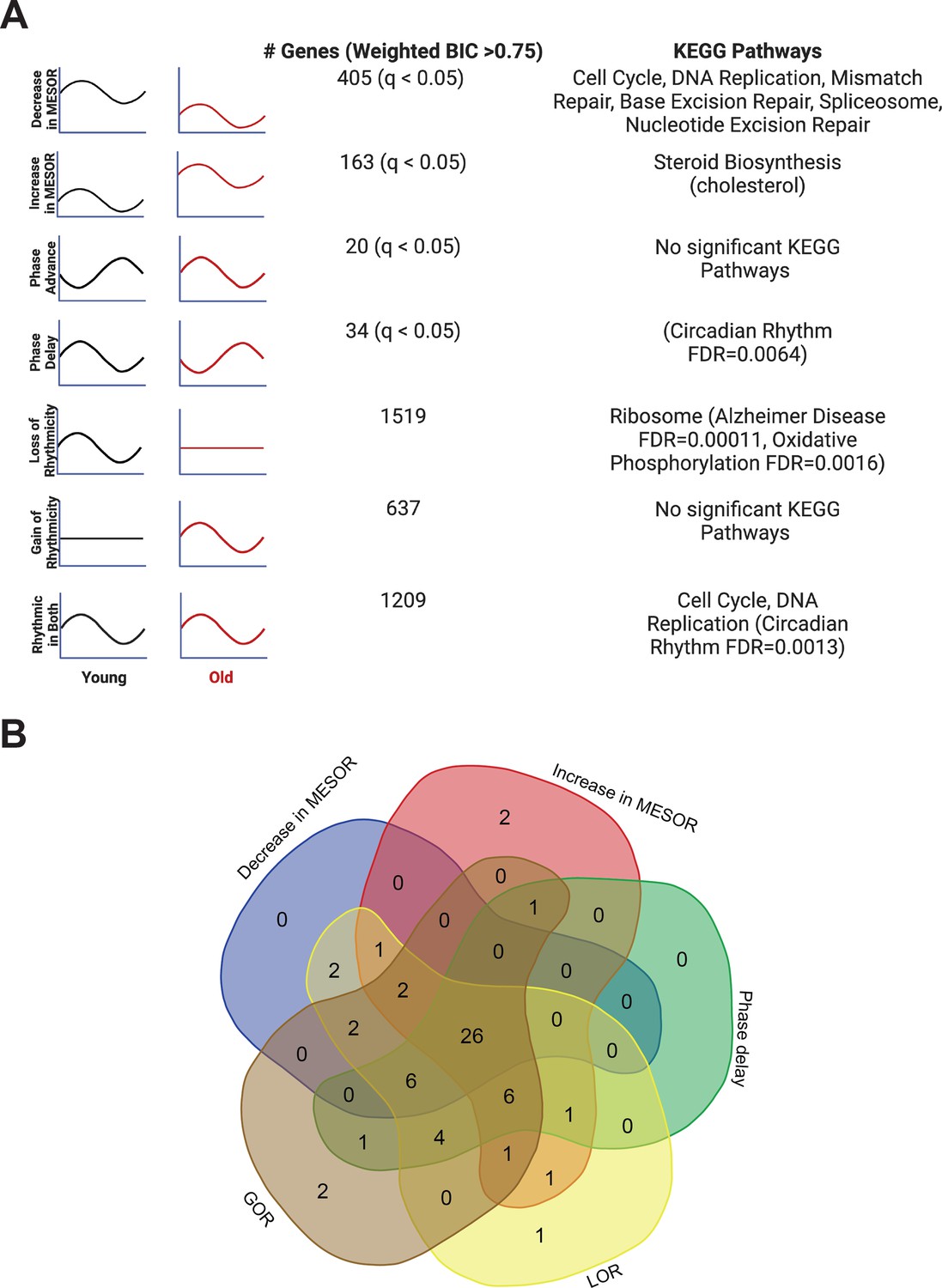
Each type of circadian change is associated with different KEGG pathways by STRING analysis, but a similar set of transcription factors identified by LISA.
Entrainment of the fibroblasts in culture with old serum significantly altered the circadian transcriptome despite use of the same cells in both young and old conditions, suggesting the aged serum itself affects the regulation of specific pathways in the cell. Significant KEGG Pathways (FDR < 1 × 10–4 unless specified) are indicated next to each category of genes. Age related pathways such as Alzheimer’s Disease/oxidative phosphorylation are associated with a loss of rhythms in the old condition. Cell cycle and DNA replication pathways remain rhythmic in the old serum condition, but cycle with a decreased MESOR. Rhythmic genes were determined using CircaCompare. Rhythmicity was determined using the weighted BIC > 0.75 criterion while p-values for difference in MESOR and phase were determined using CircaCompare (A). LinC similarity analysis (LISA), based on known transcription factor binding in fibroblasts and RNA expression, suggests that 59 total transcription factors (q<0.05) show significant changes in activity in conjunction with the following cycling phenotypes: decreased in MESOR, increased in MESOR, phase delay, gain of rhythmicity, and loss of rhythmicity as were defined above (B). Panel A was created with BioRender.com.
-
Figure 3—source data 1
Raw data from LISA transcription factor analysis based on RNA sequencing results.
- https://cdn.elifesciences.org/articles/88322/elife-88322-fig3-data1-v1.xlsx
-
Figure 3—source data 2
IPA analysis of oxidative phosphorylation genes and chromosomal replication genes that are affected by entrainment with old serum.
- https://cdn.elifesciences.org/articles/88322/elife-88322-fig3-data2-v1.xlsx
Figure 3—figure supplement 1
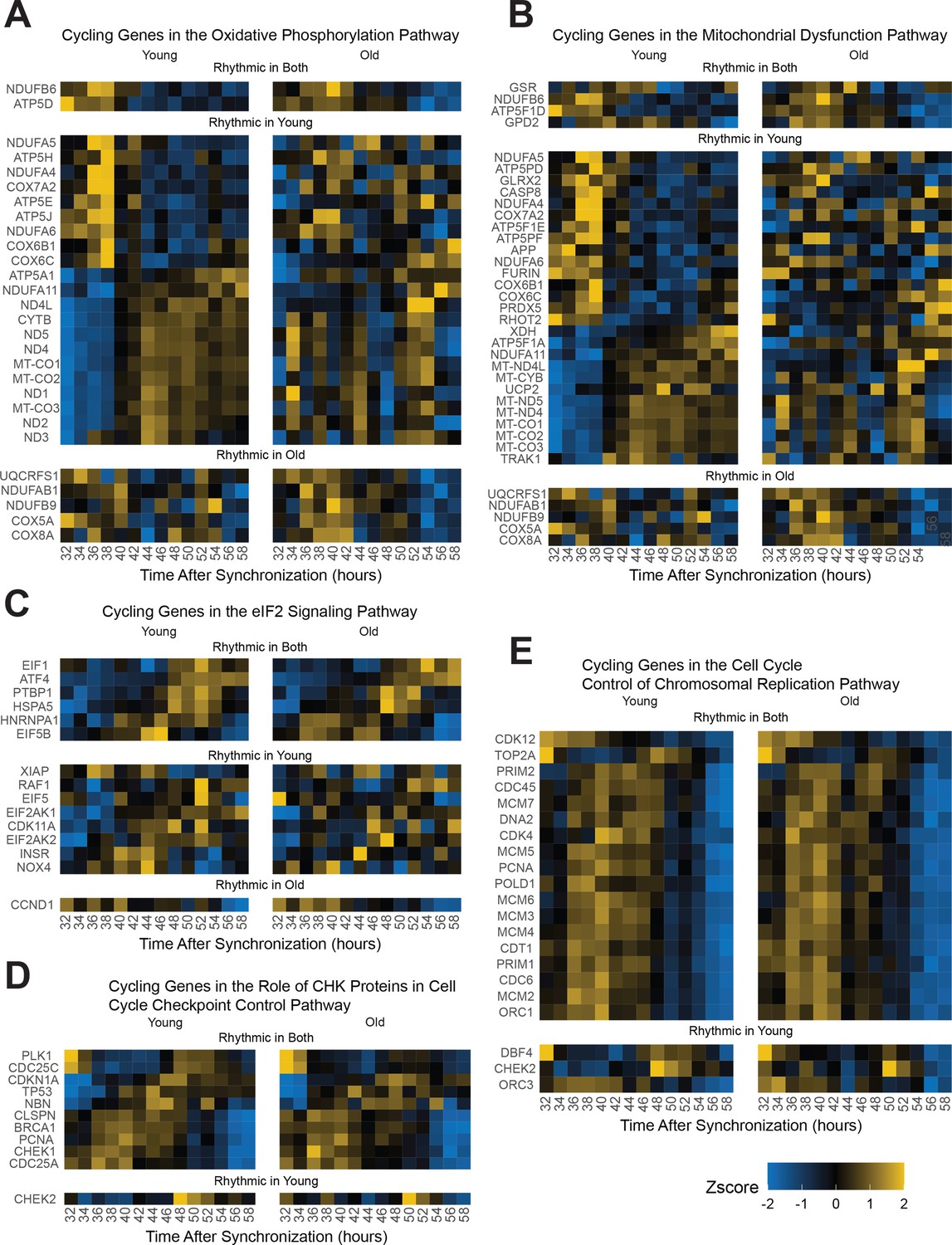
Oxidative phosphorylation genes and chromosomal replication genes are affected by entrainment with old serum.
Additional pathway analysis employing IPA further emphasizes finding with STRING analysis. Genes identified by IPA analysis to be associated with oxidative phosphorylation/mitochondrial dysfunction and eIF2 genes lose rhythmicity with age (BIC > 0.75) (A, B, C). Cell cycle checkpoint control (CHK proteins) (D) and chromosomal replication genes (E) maintain their cycling in the aged condition, which is consistent with the continued division of cells.
-
Figure 3—figure supplement 1—source data 1
Raw output from IPA analysis of genes whose transcripts cycle in at least one condition.
- https://cdn.elifesciences.org/articles/88322/elife-88322-fig3-figsupp1-data1-v1.xlsx
Figure 4 with 1 supplement
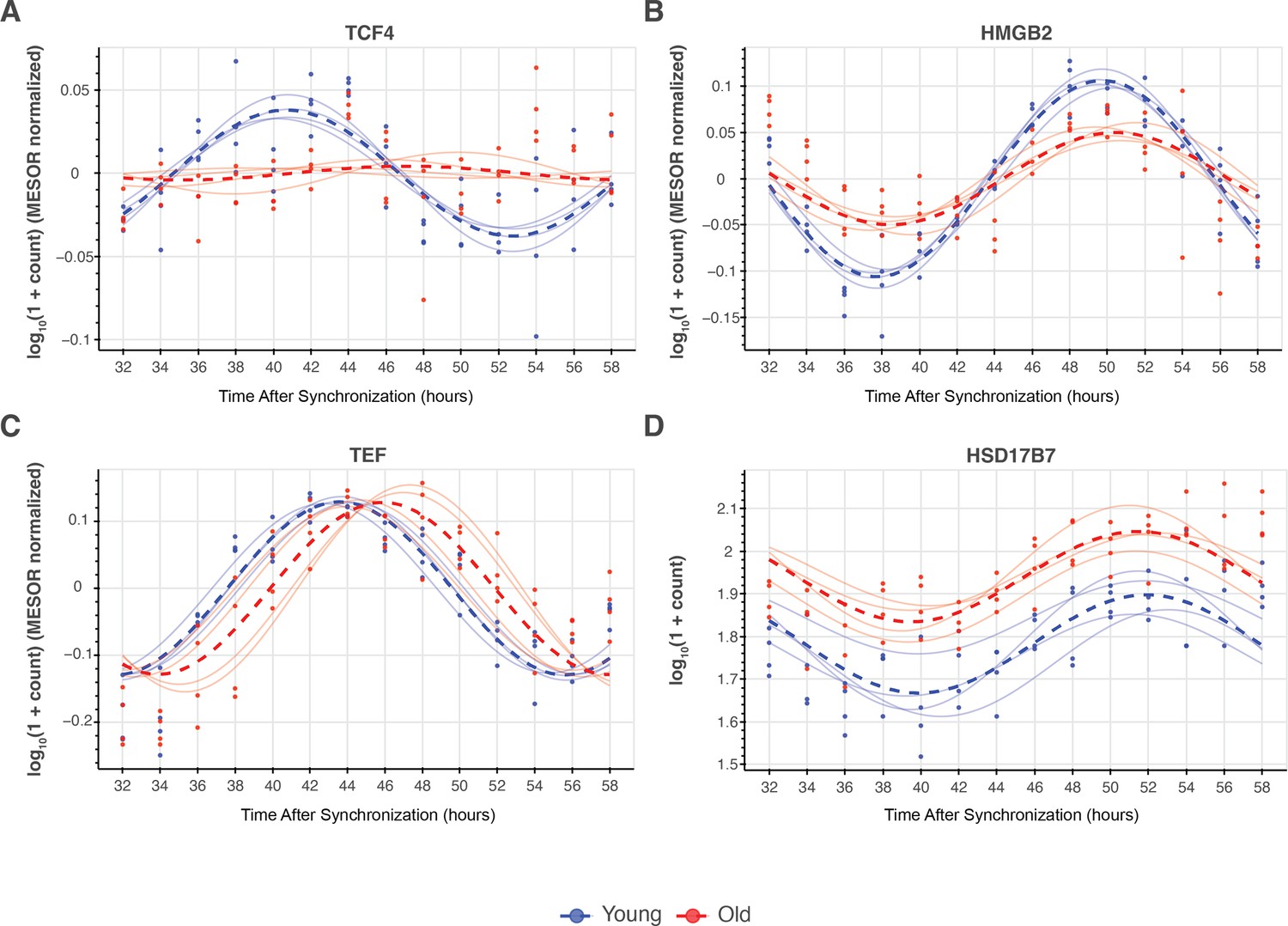
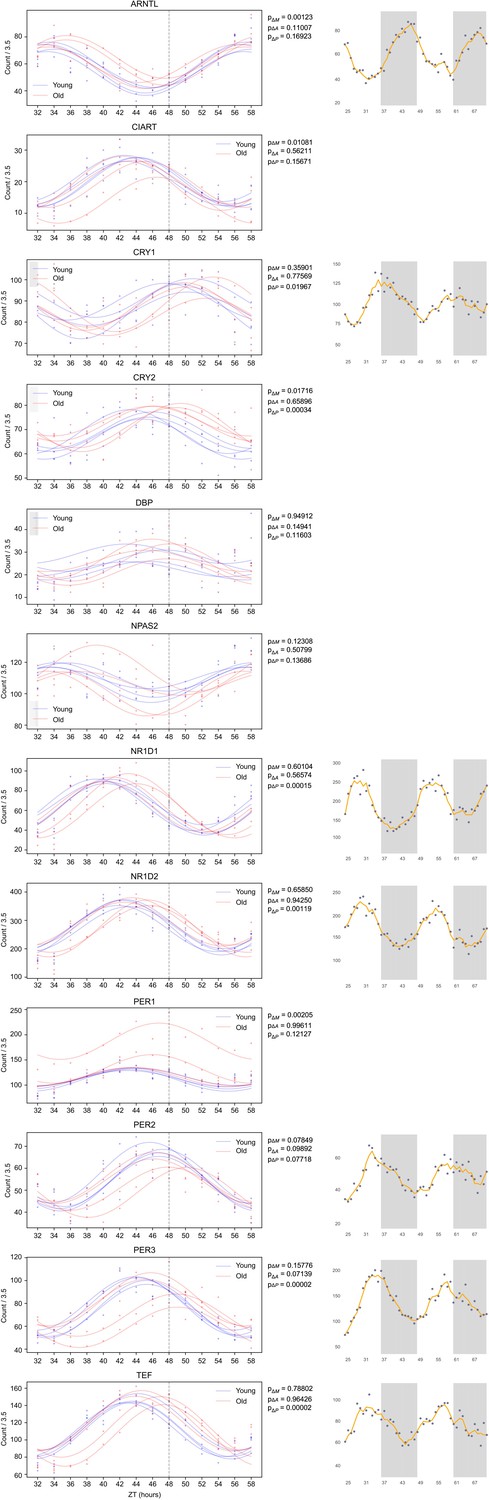
Synchronizing with old serum phase delays the expression profile of several core clock genes.
Traces of molecular clock mRNA transcripts, the average curve (bold) of results of individual sera (faded). p-Values for the difference in MESOR (P_ΔM), amplitude (P_ΔA), and phase (P_ΔP) are shown for the comparison of young and old conditions. Several clock genes are significantly phase delayed (CRY1, CRY2, NR1D1, NR1D2, PER3, TEF) in response to synchronization with old serum. While BMAL1 is not phase delayed, the MESOR significantly increases with age. N=4 subjects per timepoint for both young and old groups.
Figure 4—figure supplement 1
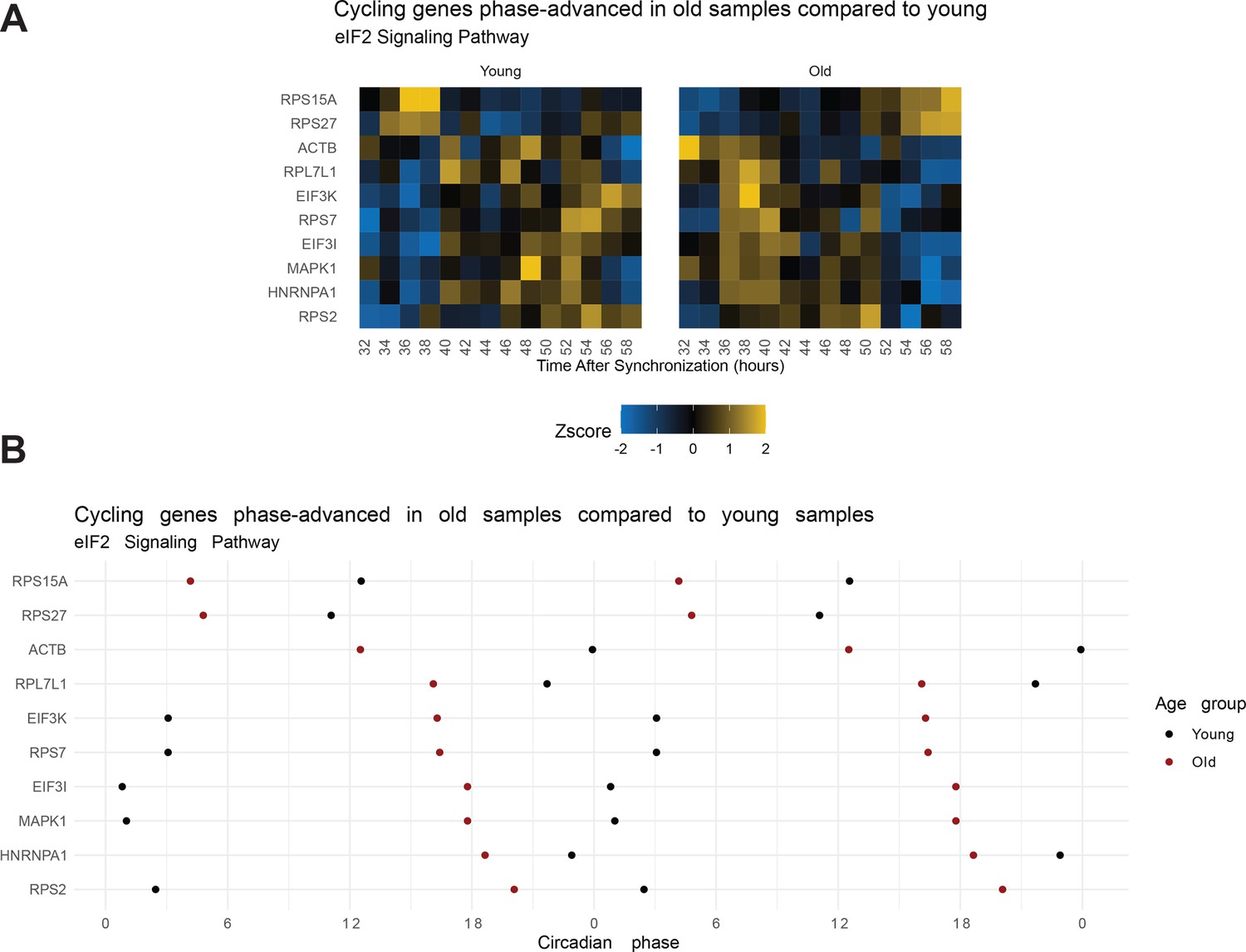
eIF2 signaling pathway genes that maintain cycling with age phase advance in the old serum condition.
From the IPA analyses, the eIF2 signaling pathway was enriched (BIC > 0.75) in two datasets: (1) genes cycling in the young sera and not the old and (2) cycling genes phase-advanced in the old sera (q<0.05). A significant number of genes that maintain rhythmicity in the old sera condition are phase advanced as visualized in the heat map (A) or plot of peak phase (B). Note, the data in the peak phase plot are double-plotted. This means the range of the x-axis is doubled and the datapoints between 0 and 24 are duplicated in the right half of the plot. Phase is a circular metric meaning a phase of 0 is equivalent to a phase of 24 (1 ⇔ 25, 2 ⇔ 26, etc). Double-plotting is used to visualize data with repeated patterns or when data straddle the 24-0 boundary. As the eIF2 pathway is involved in protein translation, this suggests that the timing of protein translation is phase advanced in the older serum and/or less rhythmic.
Author response image 1
BJ-5TA and U2OS Cells Exhibit Similar Profiles of Circadian Gene Transcription.
We compared the transcriptomic profiles of the BJ-5TA cells in young and old serum (left) to the U2OS transcriptomic data (right) available on CircaDB, a database containing profiles of several circadian reference genes in U2OS cells. This figure suggests that circadian profiles of these genes exhibit many similarities. We find that the peak to trough ratios (amplitudes) are similar for ARNTL, NR1D1, NR1D2, Per2, PER3, and that the MESORS are similar (with the exception of NR1D1 which is much lower and NR1D2 which is much higher in the BJ-5TA cells). We find that the amplitudes of CRY1 is ~25% lower and TEF is ~15% higher for the BJ5TA cells. The axis for plots on the left show counts divided by 3.5 in order to made MESORs of ARNTL similar to ease comparison.
Additional files
-
Supplementary file 1
Demographic information for subjects in the study.
Total sleep time (TST) was calculated from ActiLife 6 determined in-bed and out-of-bed times averaged across nights with available actigraphy data (≥7 nights). Old (N=8), Y-young (N=7). For the RNAseq we did a pilot experiment which had one young and one old (Supplementary file 4). The young sample didn’t have enough serum left to include that individual in the actual experiment (n=4 per group), but the old subject did. So, one old subject was used in both figures and the young were different.
- https://cdn.elifesciences.org/articles/88322/elife-88322-supp1-v1.xlsx
-
Supplementary file 2
BioPatch EKG Data Inclusion Log.
A log of which participants were included in our biopatch data based on equipment function.
- https://cdn.elifesciences.org/articles/88322/elife-88322-supp2-v1.xlsx
-
Supplementary file 3
Genes involved in the IPA cell cycle of chromosome replication pathway show decreased MESOR with age.
Table of genes that are rhythmic in both conditions with a decreased MESOR with old serum treatment compared to young serum treatment by CircaCompare. All genes are involved in the cell cycle/DNA replication pathway.
- https://cdn.elifesciences.org/articles/88322/elife-88322-supp3-v1.xlsx
-
Supplementary file 4
Genes with increased MESOR in the IPA cholesterol biosynthesis pathway.
Table of genes that are rhythmic in both conditions with a decreased MESOR with old serum treatment compared to young serum treatment by CircaCompare. All genes are involved in the cholesterol biosynthesis pathway.
- https://cdn.elifesciences.org/articles/88322/elife-88322-supp4-v1.xlsx
-
MDAR checklist
- https://cdn.elifesciences.org/articles/88322/elife-88322-mdarchecklist1-v1.docx
Download links
A two-part list of links to download the article, or parts of the article, in various formats.
Downloads (link to download the article as PDF)
Open citations (links to open the citations from this article in various online reference manager services)
Cite this article (links to download the citations from this article in formats compatible with various reference manager tools)
Evidence for a role of human blood-borne factors in mediating age-associated changes in molecular circadian rhythms
eLife 12:RP88322.
https://doi.org/10.7554/eLife.88322.3




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}