Comment on 'Parasite defensive limb movements enhance acoustic signal attraction in male little torrent frogs'
- Department of Ecology, Evolution, and Organismal Biology, Brown University, United States
- Department of Evolutionary Biology, University of Vienna, Austria
- Vienna Zoo, Austria
Abstract
Zhao et al. recently reported results which, they claim, suggest that sexual selection produces the multimodal displays seen in little torrent frogs (Amolops torrentis) by co-opting limb movements that originally evolved to support parasite defense (Zhao et al., 2022). Here, we explain why we believe this conclusion to be premature.
Introduction
Many animals communicate by performing multimodal displays that showcase vocal and gestural signals (Partan and Marler, 1999; Bro-Jørgensen, 2010; Higham and Hebets, 2013; Starnberger et al., 2014b; Mitoyen et al., 2019). Recently, Zhao et al. attempted to study how these displays might evolve, at least with respect to the process by which discrete limb movements can be incorporated into more complex signaling routines. They did this by studying little torrent frogs (Amolops torrentis), which inhabit noisy streams throughout Hainan Island in Southern China (Zhao et al., 2022). They concluded that: (i) male frogs produce a set of discrete arm and leg maneuvers to help swat away blood-sucking parasites; (ii) these same limb movements enhance the attractiveness of male calls to females. Zhao et al. then argued that natural selection for parasite-induced movements creates an opportunity for sexual selection to generate a multimodal display by integrating these movements into the species’ signaling routine. However, we argue that these conclusions are premature because they are based on misinterpretations of the study’s main results.
Results and discussion
Only “un-preferred” movements are produced around parasites
For the main conclusions of Zhao et al., 2022 to be correct, the following must be true: (i) limb movements must function to protect frogs from parasitism; (ii) this defense tactic must have emerged before the species evolved either its social limb displays or its multimodal communication strategy (True and Carroll, 2002; Borgia and Keagy, 2015; Schwark et al., 2022). However, Zhao et al. do not to provide compelling evidence for either point. For example, they report male frogs sometimes produce certain gestures when parasites land on them or when parasites fly in the frog’s “vicinity” (although this term is not defined). Moreover, they do not statistically analyze their data to assess whether frogs are more likely to produce gestures when parasites are around. We therefore ran such an analysis, and we found that only two movements —limb shaking (LSA) and wiping (W)—were more likely to occur in the presence of parasites than one might expect by chance (Figure 1). Importantly, these specific movements were not the ones that females preferred in choice tests (Figure 5C and 5D in Zhao et al., 2022). At the same time, we found that both hind foot lifting (HFL) and arm wiping (AW) were not more likely to occur in the presences of parasites (Figure 1), even though these were the two limb movements that females seemed to prefer in choice tests (Figure 5A and 5B in Zhao et al., 2022). Our results therefore suggest that parasite presence is associated with only certain limb movements that Zhao et al. studied, but none that are positively linked to female preference (but see below for concerns about female preference tests).
Figure 1
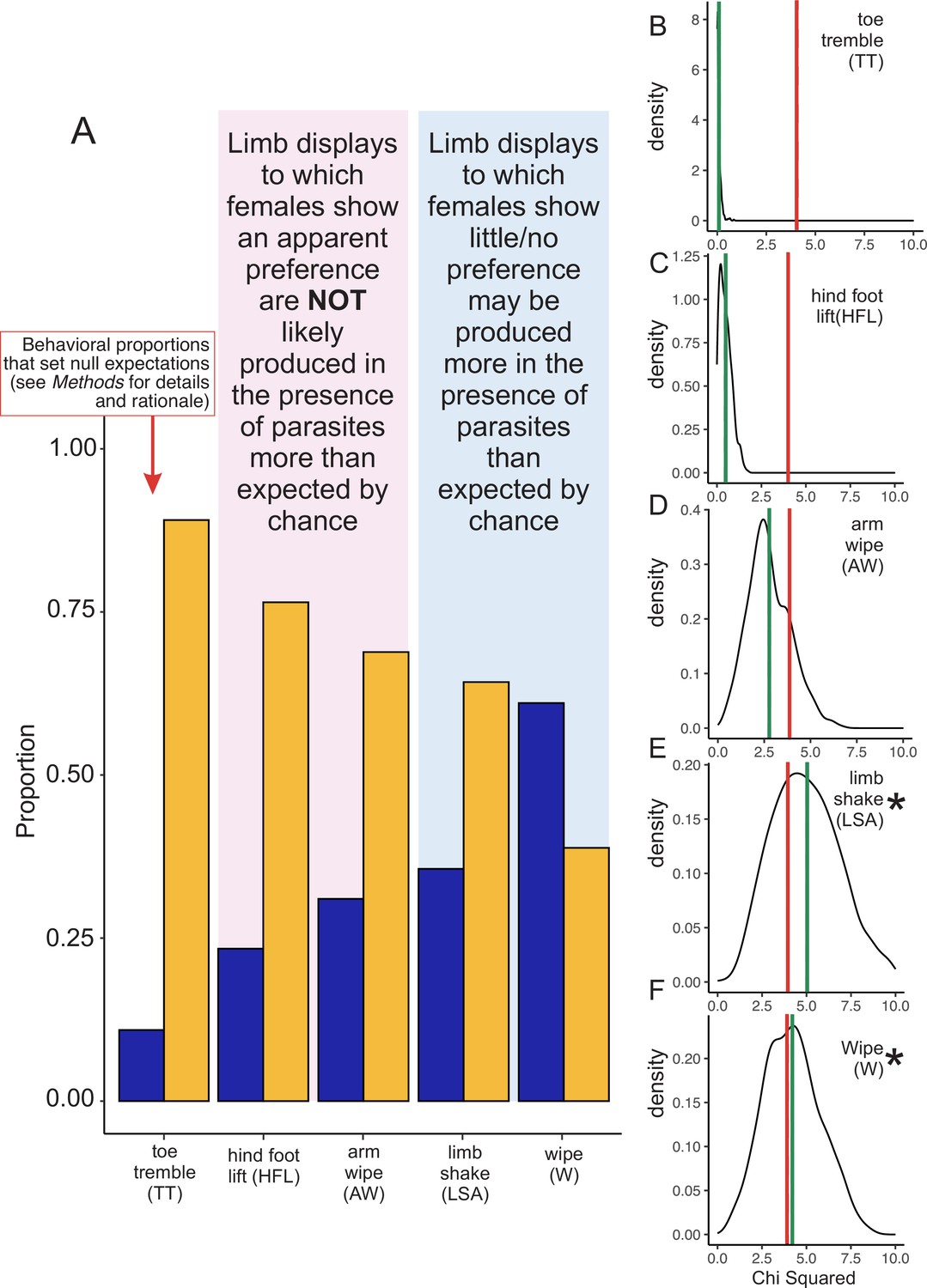
Re-analysis of whether male little torrent frogs (Amolops torrentis) produce limb displays in the presence of parasites.
(A) Proportion of different limb displays observed passively in a population of males (n=69) either in the presence of parasites (blue bars) or in the absence of parasites (orange bars). Note that these data are weighted by the number of limb movements each male produced, which were highly skewed in the original dataset. In other words, in the first analysis by Zhao et al., some males produced >90 displays, whereas other males produced zero (Zhao et al., 2022). See Methods for details about how we weighted values. For all subsequent analyses (G-tests for goodness of fit), the proportion of toe trembling produced in the presence and absence of parasites was used as the null hypothesis, setting our expectation of how often displays should be produced by chance in the presence or absence of parasites (see Methods for justification). (B–F) Density plots of the boot strapped chi-squared (χ2) statistics from the G-test of goodness of fit analysis. On the y-axis is the density of chi-squared (χ2) statistics after 1,000 iterations, and on the x-axis is the chi-squared (χ2) value. Solid green lines denote mean chi-squared statistics associated with each distribution of values, whereas solid red lines represent the cut-off for statistical significance (P<0.05) with 1 degree of freedom. If the green line falls on the right side of the red line, then the result is statistically significant (i.e., male frogs appear to perform the given display in the presence of parasites more than we might expect by chance, as determined by the null model set through toe trembling). By contrast, if the green line falls on the left side of the red line, then the result is not significant (i.e., male frogs do not perform the given display in the presence of parasites more than we might expect by chance). We found that (B) toe trembling (TT) was (as expected) not statistically significant (χ2=0.084, P=0.772), nor was (C) hind foot lifting (HFL; χ2=0.487, P=0.485) or (D) arm wiping (AW; χ2=2.772, P=0.096). Importantly, these were the behaviors the females supposedly preferred, though see the main text for a discussion of the limitations associated with this assay. We found that (E) limb shaking behavior (LSA) was statistically significant (χ2=5.0314, P=0.025, denoted with asterisk), as was (F) wiping (W) (χ2=4.212, P=0.040, denoted with asterisk). These latter two behaviors (LSA and W) were not preferred by females in the behavioral assay. Note that when comparing A to both E and F (LSA and W, respectively), the proportions in A would suggest that the effect reported in F would be more robust, compared to the effect in E. However, there were several males that did not wipe (0 values), which may have broadened the Chi Squared curve and decreased the statistical power in the analysis.
Parasitism and limb movements are correlational, and not causal
Zhao et al. also report a positive correlation between the number of parasite visits males receive and the number of limb movements males produce. They interpret these data as further support for the hypothesis that parasites are the cause of limb movements. However, correlation does not equal causation. Even if males who encountered more parasites were also more likely to have produced limb displays, this relationship does not necessarily mean that parasites directly “induced” or “evoked” this behavior, as Zhao et al. assert. Other explanations for the association include the possibility that higher quality males who display more vigorously also occupy spots along the breeding stream that contain more parasites. Micro- and macro-ecological factors that determine the abundance and distribution of blood-sucking parasites that target frogs are poorly understood (outlined recently by Virgo et al., 2022), but other work in midges implies a wide range of factors associated with the local landscape and ecology can influence their distribution and abundance (Kluiters et al., 2013; Rigot et al., 2013). Alternatively, parasites might be attracted to male calls (Bernal et al., 2006; Aihara et al., 2016; Toma et al., 2019), which males might produce more often when they are using their limbs to display during bouts of male-male competition (Grafe et al., 2012). Indeed, in both cases here, we would expect positive correlations between parasite levels and limb movements, without a causal link between the two.
Understandably, one might ask why exactly frogs would evolve limb movements like hind foot lifting (HFL) and arm wiping (AW), if they are not involved in parasite defense. This question seems even more logical given that Zhao et al. classify limbs movements produced in the absence of parasites as “spontaneous,” which implies that they are performed at random or without being triggered by an external stimulus. An alternative view, however, is that these so-called “spontaneous” limb movements are actually generated as social signals that help males compete with sexual rivals during agonistic interactions. Most frogs that use gestural signals do so for this purpose (see Table 1), and thus the behavior is assumed to evolve through intrasexual selection (Preininger et al., 2013b; Preininger et al., 2013c; Mangiamele and Fuxjager, 2018; Anderson et al., 2021a). Zhao et al. do not determine how many of the limb movements produced in the absence of parasites (e.g., “spontaneous”) were actually the result of male-male interactions, but they do indicate that little torrent frogs use these movements in such contexts.
Table 1
List of anuran species that perform limb displays or gestural signals.
For Dendrobatoidea, see Hödl and Amézquita, 2001. Note that in most cases the term courtship in the Behavioral Function column refers to instances in which females make choices about male mates, while males use gestural signals to simultaneously compete.
| Species* | Limb Signals | Sex | Behavioral Function | Evidence | Country of Origin | Ecology | Activity Pattern | Reference | |
|---|---|---|---|---|---|---|---|---|---|
| Brachychephalidae | Brachycephalus ephippium | arm waving | M | aggressive, defense | observation | Brazil | forest floor | diurnal | Pombal et al., 1994, Goutte et al., 2017 |
| B. pitanga | arm waving | M | aggressive, defense | observation | Brazil | forest floor, leaf litter | diurnal | Goutte et al., 2017 | |
| Bufonidae | Atelopus limosus | arm waving | M | context not determined | observation | Panama | stream | diurnal | Hödl and Amézquita, 2001 |
| A. varius | arm waving | M, F | aggressive, to defend sites | observation | Colombia, Costa Rica, Panama | stream | diurnal | Crump, 1988 | |
| A. zeteki | arm waving | M, F | M: agonistic, territorial vigilance, F: intersexual female-male, courtship | experimental/ mirror image | Panama | stream | diurnal | Lindquist and Hetherington, 1996, Lindquist and Hetherington, 1998 | |
| A. chiriquiensis | arm waving | M | call response, amplexus attempt | observation | Costa Rica, Panama | stream | diurnal | Lindquist and Hetherington, 1996, Lindquist and Hetherington, 1998 | |
| leg- kicking | M | during egg laying in amplexus | observation | Lindquist and Swihart, 1997 | |||||
| Leptodactylidae | Leptodactylus melanotus | foot twitching & back raise | M | aggressive | observation | Central America, Mexico | pond | Diurnal, nocturnal | Brattstrom, 1968, Gregory, 1983 |
| Crossodactylus gaudichaudii | arm waving | M | conspecific in the vicinity | observation | Brazil | stream | diurnal | Weygoldt and Potsch de Carvalho e Silva, 1992 | |
| leg stretch | M | aggressive | observation | Weygoldt and Potsch de Carvalho e Silva, 1992 | |||||
| leg lift | M | aggressive | observation | Weygoldt and Potsch de Carvalho e Silva, 1992 | |||||
| C. schmidtii | both legs kicking | M, J* | agonistic, *context not determined | observation | Brazil | stream | diurnal | Caldart et al., 2014 | |
| leg kicking | M | agonistic | observation | Caldart et al., 2014 | |||||
| toe flagging | M, F | agonistic | observation | Caldart et al., 2014 | |||||
| toe trembling | M | agonistic | observation | Caldart et al., 2014 | |||||
| limb lifting (arm & leg) | M, F, J | agonistic, M: courtship | observation | Caldart et al., 2014 | |||||
| Hylodes asper | foot flagging | M | agonistic, courtship | observation | Brazil | stream | diurnal | Haddad and Giaretta, 1999, Hartmann et al., 2005 | |
| toe movement, flagging | M | agonistic | observation | Haddad and Giaretta, 1999, Hartmann et al., 2005 | |||||
| leg stretching | M, F | M: agonistic; F: mating | observation, experimental (mirror) | Haddad and Giaretta, 1999, Hartmann et al., 2005 | |||||
| arm lifting | M | agonistic | observation | Haddad and Giaretta, 1999 | |||||
| kicking | M | aggressive | observation | Haddad and Giaretta, 1999 | |||||
| leg lifting | M | agonistic | observation | Hartmann et al., 2005 | |||||
| H. cardosoi | leg stretching (1 leg) | M | advertisement, courtship | observation | Brazil | stream | diurnal | Forti and Castanho, 2012 | |
| leg stretching (2 legs) | M | advertisement, courtship | observation | Forti and Castanho, 2012 | |||||
| limb lifting | M | advertisement, territorial | observation | Forti and Castanho, 2012 | |||||
| foot flagging | M, F | advertisement, courtship M: territorial | observation | Forti and Castanho, 2012 | |||||
| foot flagging +toe wave | M | advertisement, courtship, territorial | observation | Forti and Castanho, 2012 | |||||
| leg kicking | M | advertisement, courtship, territorial | observation | Forti and Castanho, 2012 | |||||
| H. dayctylocinus | foot flagging | M | agonistic, courtship | observation | Brazil | stream | diurnal | Narvaes and Rodrigues, 2005 | |
| toe wiggling | M | agonistic | observation | Narvaes and Rodrigues, 2005 | |||||
| leg stretching | M | agonistic | observation | Narvaes and Rodrigues, 2005 | |||||
| kicking | M | aggressive | observation | Narvaes and Rodrigues, 2005 | |||||
| arm lifting | M | context not determined | observation | Narvaes and Rodrigues, 2005 | |||||
| H. japi | toe trembling | M | agonistic, advertisement, courtship | observation | Brazil | stream | diurnal | de Sá et al., 2016 | |
| toe flagging | M | agonistic, advertisement, courtship | observation | de Sá et al., 2016 | |||||
| toes posture | M | agonistic, advertisement, courtship | observation | de Sá et al., 2016 | |||||
| foot shaking | M | agonistic, advertisement, courtship | observation | de Sá et al., 2016 | |||||
| leg stretching | M | agonistic | observation | de Sá et al., 2016 | |||||
| foot flagging | M | agonistic, advertisement, courtship | observation | de Sá et al., 2016 | |||||
| hand shaking | M | agonistic, advertisement, courtship | observation | de Sá et al., 2016 | |||||
| arm lifting | M,F | agonistic, courtship | observation | de Sá et al., 2016 | |||||
| arm waving | M,F | agonistic, courtship | observation | de Sá et al., 2016 | |||||
| H. meridionalis | toe flagging | M | agonistic | experimental | Brazil | stream | diurnal | de Sá et al., 2018, Furtado et al., 2019 | |
| toe trembling | M | agonistic | observation | de Sá et al., 2018, Furtado et al., 2019 | |||||
| toe posture | M | agonistic | observation, experimental | de Sá et al., 2018, Furtado et al., 2019 | |||||
| arm lifting | M, F | M-agonistic, F-reproductive | observation, experimental | de Sá et al., 2018, Furtado et al., 2019 | |||||
| arm waving | M, F | M-agonistic & reproductive, F-reproductive | observation, experimental | de Sá et al., 2018, Furtado et al., 2019 | |||||
| leg lifting | M, F | M-agonistic & reproductive, F-reproductive | observation, experimental | Furtado et al., 2019 | |||||
| foot flagging | M | agonistic | observation, experimental | Furtado et al., 2019 | |||||
| foot shaking | M | agonistic | observation | de Sá et al., 2018 | |||||
| both legs kicking | F | agonistic | observation | Furtado et al., 2019 | |||||
| H. nasus | toe wiggle | M | agonistic (threat signals) | observation, experimental | Brazil | stream | diurnal | Weber et al., 2004 | |
| arm waving | M | agonistic (threat signals) | observation, experimental | Weber et al., 2004 | |||||
| leg stretch | M | agonistic (threat signals) | observation, experimental | Weber et al., 2004 | |||||
| H. phyllodes | foot flagging | M | agonistic | observation, experimental | Brazil | stream | diurnal | Hartmann et al., 2005, Augusto-Alves and Toledo, 2021 | |
| leg stretching | M | agonistic, courtship | observation, experimental | Hartmann et al., 2005 | |||||
| arm lifting | M | agonistic, advertisement | observation, experimental | Hartmann et al., 2005, Augusto-Alves and Toledo, 2021 | |||||
| arm waving | M | context not determined | observation | Augusto-Alves and Toledo, 2021 | |||||
| leg lifting | M | agonistic, advertisement | observation, experimental | Hartmann et al., 2005, Augusto-Alves and Toledo, 2021 | |||||
| two limbs lifting | M | context not determined | observation | Augusto-Alves and Toledo, 2021 | |||||
| toe flagging | M | agonistic | observation, experimental | Hartmann et al., 2005, Augusto-Alves and Toledo, 2021 | |||||
| foot shaking | M | context not determined | observation | Augusto-Alves and Toledo, 2021 | |||||
| two-leg kicking | M | agonistic | observation | Augusto-Alves and Toledo, 2021 | |||||
| Myobatrachidae | Taudactylus eungellensis | leg stretching | M | context not determined | - | stream | diurnal | Hödl and Amézquita, 2001 | |
| foot flagging | M | context not determined | - | Hödl and Amézquita, 2001 | |||||
| Hylidae | Boana albomarginata | Limb lifting | M | agonistic | experimental (mirror) | Brazil | pond margins vegetation | nocturnal | Hartmann et al., 2005, Furtado and Nomura, 2014 |
| (Hypsiboas albomarginatus) | face wiping | M | agonistic | experimental (mirror) | Furtado and Nomura, 2014 | ||||
| (Hyla albormarginata) | toe trembling | M | agonistic | experimental (mirror) | Hartmann et al., 2005, Furtado and Nomura, 2014 | ||||
| leg kicking | M | agonistic | experimental (mirror) | Hartmann et al., 2005, Furtado and Nomura, 2014 | |||||
| B. raniceps | limb lifting | M | agonistic | experimental (mirror) | Brazil | ponds or wetlands | nocturnal | Furtado et al., 2017 | |
| (Hypsiboas raniceps) | toe/finger trembling | M | agonistic | experimental (mirror) | stream | nocturnal | Furtado et al., 2017 | ||
| Litoria cooloolensis | foot flagging | M | agonistic | observation | Australia | tree | nocturnal | Meyer et al., 2012 | |
| L. genimaculata | foot flagging | M | agonistic | observation | Australia | stream | nocturnal | Richards and James, 1992 | |
| L. iris | leg flicking | M | call response | observation | Papua New Guinea | stream | crepuscular | Meyer et al., 2012 | |
| L. nannotis | foot flagging | M | agonistic | observation | Australia | stream | nocturnal | Richards and James, 1992 | |
| arm waving | M | agonistic | observation | nocturnal | Richards and James, 1992 | ||||
| L. pearsoniana | hand waving | M | agonistic | observation | Australia | stream | nocturnal | Meyer et al., 2012 | |
| leg flicking | M | agonistic | observation | Meyer et al., 2012 | |||||
| L.rheocola | leg stretching | M | agonistic | observation | Australia | stream | nocturnal | Richards and James, 1992 | |
| arm waving | M | agonistic | observation | Richards and James, 1992 | |||||
| L. fallax | foot flagging | M | agonistic | observation | Australia | pond | nocturnal | Meyer et al., 2012 | |
| foot flickering | M | agonistic | observation | Meyer et al., 2012 | |||||
| kicking | M | aggressive | observation | Meyer et al., 2012 | |||||
| Lysapsus limellum | Limb lifting | M | agonistic | experimental (mirror) | Brazil | lentic water bodies | nocturnal | Furtado et al., 2017 | |
| Dendropsophus nanus | Limb lifting | M | agonistic | experimental (mirror) | Brazil | ponds | nocturnal | Furtado et al., 2017 | |
| Dendropsophus parviceps | foot flagging | M | agonistic | observation | Venezuela | streamside ponds | nocturnal | Amézquita and Hödl, 2004 | |
| Hyla parviceps | arm waving | M | agonistic | observation | Amézquita and Hödl, 2004 | ||||
| Hyla sp. (aff. ehrhardti) | body wiping (foot) | courtship | observation | Brazil | forest, bromeliads | nocturnal | Hartmann et al., 2005 | ||
| face wiping (arm) | M, F | courtship | observation | Hartmann et al., 2005 | |||||
| foot flagging | M | courtship (far from females) | observation | Hartmann et al., 2005 | |||||
| limb lifting (arm +leg) | M | courtship | observation | Hartmann et al., 2005 | |||||
| Phyllomedusa boliviana | foot flagging | M | aggressive | observation | Bolivia | pond | nocturnal | Jansen and Kohler, 2008 | |
| leg lifting | M | aggressive | observation | Jansen and Kohler, 2008 | |||||
| leg stretching | M | aggressive | observation | Jansen and Kohler, 2008 | |||||
| P. burmeisteri | leg stretching | M | agonistic | observation | Brazil | pond | nocturnal | Abrunhosa and Wogel, 2004 | |
| kicking | M | aggressive | observation | Abrunhosa and Wogel, 2004 | |||||
| P. sauvagii | foot flagging | M | territorial | observation | Argentina, Bolivia, Paraguay, Brazil | pond | nocturnal | Halloy and Espinoza, 2000 | |
| Scinax eurydice | leg kicking | M | 2 males far from each other | observation | Brazil | pond (rainy season) | nocturnal | Hartmann et al., 2005 | |
| limb lifting (arm +leg) | M | 2 males far from each other | observation | Hartmann et al., 2005 | |||||
| Centrolenidae | Vitreorana uranoscopa | limb lifting (arm +leg) | M | agonistic, spontaneous (no other individual present) | observation | Brazil | nocturnal | Hartmann et al., 2005 | |
| (Hyalinobatrachium uranoscopum) | |||||||||
| Ranidae | Pulchrana (Rana) baramica | toe waving | M | attract prey | observation | Singapore | forest | Grafe, 2008 | |
| Staurois latopalmatus | arm waving | M | agonistic | observation | Borneo | stream | diurnal | Preininger et al., 2009 | |
| foot flagging | M | agonistic | observation | Preininger et al., 2009 | |||||
| S. guttatus | foot flagging | M, F | agonistic | F-experimental, M-observation | stream | Grafe and Wanger, 2007, Preininger et al., 2016 | |||
| leg drumming | M | context not determined | observation | Grafe and Wanger, 2007 | |||||
| foot raising | M | courtship | observation | Grafe and Wanger, 2007 | |||||
| arm waving | M | agonistic | observation | Grafe and Wanger, 2007 | |||||
| S. parvus | foot flagging | M, J | agonistic | observation, experimental | Borneo | stream | diurnal | Grafe et al., 2012, Preininger et al., 2012, Preininger et al., 2013b | |
| foot lifting (tap) | M | agonistic | observation, experimental | Grafe et al., 2012, Preininger et al., 2013b | |||||
| Micrixalidae | Micrixalus candidus | foot lifting | M | agonistic | observation | India | stream | diurnal | Preininger and Fuxjager, pers. observation |
| foot stretching | M | agonistic | observation | Preininger and Fuxjager, pers. observation | |||||
| foot flagging | M | agonistic | observation | Preininger and Fuxjager, pers. observation | |||||
| M. elegans | foot lifting | M | agonistic | observation | India | stream | diurnal | Preininger and Fuxjager, pers. observation | |
| foot stretching | M | agonistic | observation | Preininger and Fuxjager, pers. observation | |||||
| foot flagging | M | agonistic | observation | Preininger and Fuxjager, pers. observation | |||||
| M. kottigeharensis | foot lifting | M | agonistic | observation | India | stream | diurnal | Preininger et al., 2013c, Anderson et al., 2021b, Anderson et al., 2021d | |
| foot stretching | M | agonistic | observation | Preininger et al., 2013b, Preininger et al., 2013c | |||||
| foot flagging | M | agonistic | observation | Preininger and Fuxjager, pers. observation | |||||
| toe wiggling | M | agonistic | observation | Preininger and Fuxjager, pers. observation | |||||
| kicking | M | aggressive | observation | Preininger et al., 2013c | |||||
| M. niluvasei | foot lifting | M | agonistic | observation | India | stream | Anderson et al., 2021d, Preininger and Fuxjager, pers. observation | ||
| foot stretching | M | agonistic | observation | Preininger and Fuxjager, pers. observation | |||||
| foot flagging | M | agonistic | observation | Preininger and Fuxjager, pers. observation | |||||
| kicking | M | aggressive | observation | Preininger and Fuxjager, pers. observation | |||||
| M. saxicola | foot lifting | M | agonistic | observation | India | stream | Anderson et al., 2021d, Preininger and Fuxjager, pers. observation | ||
| foot stretching | M | agonistic | observation | Preininger and Fuxjager, pers. observation | |||||
| foot flagging | M | agonistic | observation | Preininger and Fuxjager, pers. observation | |||||
| toe wiggling | M | agonistic | observation | Preininger and Fuxjager, pers. observation | |||||
| kicking | M | aggressive | observation | Preininger and Fuxjager, pers. observation | |||||
| M. specca | foot lifting | M | agonistic | observation | India | stream | Preininger and Fuxjager, pers. observation | ||
| foot flagging | M | agonistic | observation | Preininger and Fuxjager, pers. observation | |||||
| M. uttaraghati | foot lifting | M | agonistic | observation | India | stream | Preininger and Fuxjager, pers. observation | ||
| foot stretching | M | agonistic | observation | Preininger and Fuxjager, pers. observation | |||||
| foot flagging | M | agonistic | observation | Preininger and Fuxjager, pers. observation | |||||
| toe wiggling | M | agonistic | observation | Preininger and Fuxjager, pers. observation | |||||
| Rhacophoridae | Buergeria japonica | leg-stretches | M | agonistic male-male interaction | observation | Japan | aquatic and terrestrial | Anderson et al., 2021c | |
| B. otai | foot-flagging | M | agonistic male-male interaction | observation | Taiwan | stream | Yang, 2022 | ||
| Theloderma bambusicolum | foot-flagging | M | territorial behavior | observation | Vietnam | dense bushes | Orlov et al., 2012 |
-
*
Species names in parentheses represent former names used in original publication.
-
M=male; F=female; J=juvenile.
Limitations to the female preference tests
Zhao et al. also conduct experiments that examine whether females prefer to associate with males that produce supposed “parasite-induced” limb movements while calling. In theory, results from this study should provide the rationale for the hypothesis that sexual selection by female choice co-opts leg movements into reproductive displays. Yet, as we indicate above, this idea runs counter to many studies that suggest that gestural displays in frogs mediate agonistic encounters among males (Table 1). To our knowledge, there are currently no studies that clearly and definitively show that male frogs use the same limb movements described by Zhao et al. to attract female mates. There is certainly some observational evidence for visual displays employed during courtship, but such data are relatively rare and functionally ambiguous (examples: de Sá et al., 2018 has n=3 courtship interactions; Furtado et al., 2019 has n=1 courtship interaction). To this end, Zhao et al. only report four male-female interactions across two breeding seasons, and during these interactions males don’t produce any of the limb displays that are purported to be linked with parasite defense. Furthermore, when working in the field with torrent frogs, one must recognize that it is nearly impossible to distinguish male gestural displays directed to other males from those directed at females (see Table 1 and most “courtship” interactions listed therein). This is because males in the area will trigger these behaviors from each other, even as females approach (Preininger and Fuxjager, personal observations; Zhao et al., 2022).
Still, Zhao et al. attempt to test female preference for male limb movements by presenting females with video stimuli of males that were calling and either producing limb movements or not. However, these video stimuli are not ecologically relevant to female frogs. This is because each stimulus was manually altered to include a standardized audio channel, such that the male in the video would be perceived to have called without inflating its vocal sac. Free-living females do not naturally encounter such stimuli, particularly when they assess males by looking at them head-on (as females do in this experiment). Zhao et al. indicate that they designed the stimuli this way because they were afraid the effect of vocal sac inflation would mask any effect of limb movement on female preference. Vocal sac inflation has a powerful effect on sexual attractiveness and mate choice in frogs (reviewed by Starnberger et al., 2014a), including in little torrent frogs (Zhao et al., 2021). Importantly, if vocal sac inflation does mask effects of limb movements on female preference, then selection should not strongly favor the co-option of these movements into the display. We suspect that females showed a preference for males that produced HFL and AW movements because they were the closest resemblance of “fixed” vocal sac inflations, particularly when the alternative stimulus included calls without vocal sac inflations (Rosenthal et al., 2004; Narins et al., 2005; Taylor et al., 2008; Gomez et al., 2011; Preininger et al., 2013a). Visual and acoustic components might differ in context and dominance, but nevertheless strongly modulate mate choice (Taylor et al., 2011). One might argue against our point by saying that females can in fact observe males producing limb movements and calls without seeing vocal sac inflation, such as when females see males from behind. However, such visual perspectives of the male were not incorporated into the experimental design, and thus the current study cannot reveal how females would respond to seeing males perform limb movements from such alternate angles.
Conclusions
Here, we highlight concerns about a study by Zhao et al. that tried to explain the origins of multimodal display behavior in little torrent frogs (Zhao et al., 2022). By reanalyzing data from this study, we show that only certain limb movements are potentially performed more in the presence of parasites, and these are not the movements that females seem to prefer. The study by Zhao et al. also over-interprets correlational evidence to propose that limb movements evolved to avoid parasite attacks. Finally, Zhao et al. cannot determine whether limb movements are functionally significant during male-female interactions because female preference experiments were limited with respect to their ethological relevance.
We also have other concerns about this study. For example, data videos and drawings of limb movements are ambiguous and unclear (e.g., parasites are unclear in Video 1; gesture illustrations in Figure 1E and C show mirror images of the same movements), and there are no data showing how frequently frogs use limb movements to physically wipe away parasites, or whether frogs ever experience parasites in their “vicinity” without producing limb movements. It is also unclear why preference tests were carried out at night, which creates a temporal mismatch with day-recorded video stimuli. Nonetheless, as biologists who study gestural signals in frogs, we remain open to the possibility that visual displays might arise through the co-option of adaptive movements that are unrelated to communication. Similarly, we recognize that the role of female choice in the evolution of frog limb displays is poorly understood and merits further investigation. However, studies exploring these topics should be carried out using approaches that are clear and replicable, so that we can draw lasting conclusions.
Materials and methods
We used data from the original study (Table S1 in Zhao et al., 2022) to statistically test whether male frogs were more likely to produce the various limb movements when parasites were around than one would otherwise expect by chance. We reasoned that this analysis would help us understand whether behaviors that were more closely aligned with parasite presence were also associated with female preference tests. (Please see above for a discussion of the limitations associated with preference tests).
We ran all statistical analyses in R Studio (https://www.rstudio.com), an integrated environment for R 4.13 (https://www.r-project.org). For data preprocessing, we noted that Zhao et al. did not account for the drastic differences in number of behaviors produced by each frog. This oversight can lead to certain individuals in the population having an outsized effect on statistical outcomes. For example, a frog that produced ≈90 limb movements in the absence of parasites and 10 limb movements in the presence of parasites was compared to another frog that produced 10 limb movements in absence of parasites and 1 limb movement in the presence of parasites. The proportion of behaviors that these individuals produced in each context is the same, but the absolute total number of these behaviors is quite different; as a result, if raw values of behavior are compared between the groups (absence of parasites vs. presence of parasites), then the first frog will have a more robust impact than the second frog. Weighting values can be an important way to avoid such effects, and so we adopted this approach. We weighted following Garamszegi, 2014, where each display count, X, was multiplied by the inverse of the sum count of X for the given individual.
Next, to test how limb movements might correspond to the presence of parasites, we used a G-test (for goodness of fit) to statistically compare the proportion of limb movements produced in the absence of parasites (i.e., called “spontaneous” limb movements, see main text) and the proportion of limb movements produced in the presence of parasites. This test assumes independence between the proportions. To meet this assumption, we randomly sampled 35 individuals from the data set, and noted the total number of “spontaneous” limb movements these individuals produced. We then took the remaining 34 individuals from the data set and recorded only the total number of limb movements produced in the presence of parasites. We repeated this process 1,000 times, always resampling the dataset with replacement. In each case, we employed the g.test function from the AMR package to calculate a Chi Squared (χ2) test statistic, which produced a distribution of statistic values. We used the mean χ2 statistic associated with each limb movement to compute a corresponding p value. Importantly, these models were calculated using a null distribution that was determined by the level of toe trembling behavior in the absence (89%) and presence of parasites (11%). Past studies, including some that Zhao et al. cite (such as Hödl and Amézquita, 2001), show that toe trembling is not a parasite defense behavior; rather, it is commonly used either as a social signal (Lindquist and Hetherington, 1996; Rojas and Pašukonis, 2019) or as a feeding/hunger signal (Grafe, 2008; Hagman and Shine, 2008; Sloggett and Zeilstra, 2008; McFadden et al., 2010; Claessens et al., 2020). Either way, toe trembling provides a nice statistical heuristic to anchor our a priori expectations of how many of these limb displays should be produced when parasites are not around vs. when they are around. Accordingly, if the proportion of limb displays differed significantly from this expectation, then we could conclude that the given behavior was produced more often in the presence of parasites than expected by chance. By contrast, if the proportion of limb displays did not differ significantly from our null expectation based on toe trembling, then we cannot reject the null hypothesis.
Data availability
Figure 1 source data are included with original manuscript (Supplementary file 1) on which we are commenting.
-
Dryad Digital RepositoryThe data of parasite-induced and spontaneous displays in each limb movement for calling males, silent males and males that have females nearby.https://doi.org/10.5061/dryad.f1vhhmgzg
References
-
How, when, and where to perform visual displays? The case of the Amazonian frog Hyla parvicepsHerpetologica 60:20–29.
-
Testosterone amplifies the negative valence of an agonistic gestural display by exploiting receiver perceptual biasProceedings of the Royal Society B: Biological Sciences 288:20211848.https://doi.org/10.1098/rspb.2021.1848
-
Insight into the evolution of anuran foot flag displays: A comparative study of color and kinematicsIchthyology & Herpetology 109:1047–1059.https://doi.org/10.1643/h2020160
-
A common endocrine signature marks the convergent evolution of an elaborate dance display in frogsThe American Naturalist 198:522–539.https://doi.org/10.1086/716213
-
Cognitively driven co‐option and the evolution of complex sexual displays in bowerbirdsAnimal Signaling and Function 1:75–109.https://doi.org/10.1002/9781118966624
-
Dynamics of multiple signalling systems: Animal communication in a world in fluxTrends in Ecology & Evolution 25:292–300.https://doi.org/10.1016/j.tree.2009.11.003
-
Notes on agonistic communication by the Neotropical torrent frog Hylodes meridionalis (Hylodidae)Herpetology Notes 11:919–923.
-
Behavioural repertoire and a new geographical record of the torrent frog Hylodes cardosoi (Anura: Hylodidae)Herpetological Bulletin 121:17–22.
-
In front of a mirror: visual displays may not be aggressive signals in nocturnal tree frogsJournal of Natural History 51:443–454.https://doi.org/10.1080/00222933.2016.1262078
-
Toe waving in the brown marsh frog Rana baramica: pedal luring to attract prey?Scientia Bruneiana 9:3–5.
-
Visual and acoustic communication in the Brazilian torrent frog, Hylodes asper (Anura: Leptodactylidae)Herpetologica 55:324–333.
-
Territorial encounters and threat displays in the neotropical frog Phyllomedusa sauvagii (Anura: Hylidae)Herpetological Natural History 7:175–180.
-
Visual communication in Brazilian species of anurans from the Atlantic forestJournal of Natural History 39:1675–1685.https://doi.org/10.1080/00222930400008744
-
An introduction to multimodal communicationBehavioral Ecology and Sociobiology 67:1381–1388.https://doi.org/10.1007/s00265-013-1590-x
-
BookVisual signaling in Anuran AmphibiansIn: Ryan MJ, editors. Anuran Communication. Smithsonian Institute Press. pp. 121–141.
-
Intraspecific combat behavior of Phyllomedusa boliviana (Anura: Hylidae) and the possible origin of visual signaling in nocturnal treefrogsHerpetological Review 39:290.
-
Modelling the spatial distribution of Culicoides biting midges at the local scaleJournal of Applied Ecology 50:232–242.https://doi.org/10.1111/1365-2664.12030
-
Atelopus chiriquiensis (Chiriqui Harlequin Frog). Mating behaviour and egg-layingHerpetological Review 3:145.
-
Insight into the neuroendocrine basis of signal evolution: A case study in foot-flagging frogsJournal of Comparative Physiology A 204:61–70.https://doi.org/10.1007/s00359-017-1218-0
-
Toe-twitching during feeding in the Australian myobatrachid frog, Pseudophryne corroboreeHerpetological Review 41:153–154.
-
Taxonomic notes on rhacophorid frogs (Rhacophorinae: Rhacophoridae: Anura) of southern part of Annamite Mountains (Truong Son, Vietnam), with description of three new speciesRussian Journal of Herpetology 19:23–64.
-
The conservation breeding of two foot-flagging frog species from Borneo, Staurois parvus and Staurois guttatusAmphibian and Reptile Conservation 5:45–56.
-
Multimodal signaling in the Small Torrent Frog (Micrixalus saxicola) in a complex acoustic environmentBehavioral Ecology and Sociobiology 67:1449–1456.https://doi.org/10.1007/s00265-013-1489-6
-
Getting a kick out of it: multimodal signalling during male–male encounters in the foot-flagging frog Micrixalus aff. saxicola from the Western Ghats of IndiaCurrent Science 105:1735–1740.
-
Comparison of female and male vocalisation and larynx morphology in the size dimorphic foot-flagging frog species Staurois guttatusThe Herpetological Journal 26:187–197.
-
Foot-flagging displays of some Australian frogsMemoirs of the Queensland Museum 32:302.
-
From uni- to multimodality: towards an integrative view on anuran communicationJournal of Comparative Physiology. A, Neuroethology, Sensory, Neural, and Behavioral Physiology 200:777–787.https://doi.org/10.1007/s00359-014-0923-1
-
The anuran vocal sac: a tool for multimodal signallingAnimal Behaviour 97:281–288.https://doi.org/10.1016/j.anbehav.2014.07.027
-
Multimodal signal variation in space and time: how important is matching a signal with its signaler?Journal of Experimental Biology 214:815–820.https://doi.org/10.1242/jeb.043638
-
Gene co-option in physiological and morphological evolutionAnnual Review of Cell and Developmental Biology 18:53–80.https://doi.org/10.1146/annurev.cellbio.18.020402.140619
-
Behavioral and neurogenomic responses to acoustic and visual sexual cues are correlated in female torrent frogsAsian Herpetological Research 12:88–99.https://doi.org/10.16373/j.cnki.ahr.200063
Article and author information
Author details
Matthew J Fuxjager

Funding
National Science Foundation (OISE-1952542)
- Matthew J Fuxjager
Vienna Zoo
- Doris Preininger
The funders had no role in study design, data collection and interpretation, or the decision to submit the work for publication.
Acknowledgements
We thank Nick Antonson, Nicole Moody, and Sofia Piggott for helpful discussions about this paper.
Copyright
© 2023, Anderson et al.
This article is distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use and redistribution provided that the original author and source are credited.
Metrics
-
- 350
- views
-
- 34
- downloads
-
- 3
- citations
Views, downloads and citations are aggregated across all versions of this paper published by eLife.
Download links
A two-part list of links to download the article, or parts of the article, in various formats.
Downloads (link to download the article as PDF)
Open citations (links to open the citations from this article in various online reference manager services)
Cite this article (links to download the citations from this article in formats compatible with various reference manager tools)
Comment on 'Parasite defensive limb movements enhance acoustic signal attraction in male little torrent frogs'
eLife 12:e89134.
https://doi.org/10.7554/eLife.89134
Further reading
-
- Ecology
- Evolutionary Biology
Many animals rely on complex signals that target multiple senses to attract mates and repel rivals. These multimodal displays can however also attract unintended receivers, which can be an important driver of signal complexity. Despite being taxonomically widespread, we often lack insight into how multimodal signals evolve from unimodal signals and in particular what roles unintended eavesdroppers play. Here, we assess whether the physical movements of parasite defense behavior increase the complexity and attractiveness of an acoustic sexual signal in the little torrent frog (Amolops torrentis). Calling males of this species often display limb movements in order to defend against blood-sucking parasites such as frog-biting midges that eavesdrop on their acoustic signal. Through mate choice tests we show that some of these midge-evoked movements influence female preference for acoustic signals. Our data suggest that midge-induced movements may be incorporated into a sexual display, targeting both hearing and vision in the intended receiver. Females may play an important role in incorporating these multiple components because they prefer signals which combine multiple modalities. Our results thus help to understand the relationship between natural and sexual selection pressure operating on signalers and how in turn this may influence multimodal signal evolution.
-
- Ecology
Global change is causing unprecedented degradation of the Earth’s biological systems and thus undermining human prosperity. Past practices have focused either on monitoring biodiversity decline or mitigating ecosystem services degradation. Missing, but critically needed, are management approaches that monitor and restore species interaction networks, thus bridging existing practices. Our overall aim here is to lay the foundations of a framework for developing network management, defined here as the study, monitoring, and management of species interaction networks. We review theory and empirical evidence demonstrating the importance of species interaction networks for the provisioning of ecosystem services, how human impacts on those networks lead to network rewiring that underlies ecosystem service degradation, and then turn to case studies showing how network management has effectively mitigated such effects or aided in network restoration. We also examine how emerging technologies for data acquisition and analysis are providing new opportunities for monitoring species interactions and discuss the opportunities and challenges of developing effective network management. In summary, we propose that network management provides key mechanistic knowledge on ecosystem degradation that links species- to ecosystem-level responses to global change, and that emerging technological tools offer the opportunity to accelerate its widespread adoption.




{kind=link}