Exposure to live saprophytic Leptospira before challenge with a pathogenic serovar prevents severe leptospirosis and promotes kidney homeostasis
- Department of Microbiology, Immunology and Biochemistry, University of Tennessee Health Science Center, United States
eLife assessment
This important study contributes to our understanding on how prior exposure to a non-pathogenic Leptospira strain could prime the host to prevent severe leptospirosis following infection with a pathogenic strain. The work described is solid and broadly supports the claims, with minor weaknesses that could be addressed in future studies. The work will be of interest to scientists interested in host-pathogen interactions and leptospirosis.
https://doi.org/10.7554/eLife.96470.3.sa0Significance of the findings:
Important: Findings that have theoretical or practical implications beyond a single subfield
- Landmark
- Fundamental
- Important
- Valuable
- Useful
Strength of evidence:
Solid: Methods, data and analyses broadly support the claims with only minor weaknesses
- Exceptional
- Compelling
- Convincing
- Solid
- Incomplete
- Inadequate
During the peer-review process the editor and reviewers write an eLife Assessment that summarises the significance of the findings reported in the article (on a scale ranging from landmark to useful) and the strength of the evidence (on a scale ranging from exceptional to inadequate). Learn more about eLife Assessments
Abstract
Previous studies demonstrated that Leptospira biflexa, a saprophytic species, triggers innate immune responses in the host during early infection. This raised the question of whether these responses could suppress a subsequent challenge with pathogenic Leptospira. We inoculated C3H/HeJ mice with a single or a double dose of L. biflexa before challenge with a pathogenic serovar, Leptospira interrogans serovar Copenhageni FioCruz (LIC). Pre-challenge exposure to L. biflexa did not prevent LIC dissemination and colonization of the kidney. However, it rescued weight loss and mouse survival thereby mitigating disease severity. Unexpectedly, there was correlation between rescue of overall health (weight gain, higher survival, lower kidney fibrosis marker ColA1) and higher shedding of LIC in urine. This stood in contrast to the L. biflexa unexposed LIC challenged control. Immune responses were dominated by increased frequency of effector T helper (CD4+) cells in spleen, as well as significant increases in serologic IgG2a. Our findings suggest that exposure to live saprophytic Leptospira primes the host to develop Th1 biased immune responses that prevent severe disease induced by a subsequent challenge with a pathogenic species. Thus, mice exposed to live saprophytic Leptospira before facing a pathogenic serovar may withstand infection with far better outcomes. Furthermore, a status of homeostasis may have been reached after kidney colonization that helps LIC complete its enzootic cycle.
Introduction
Leptospirosis, a neglected re-emerging enzootic spirochetal disease, affects millions of people worldwide causing an overall mortality rate of 65,000 per year (Costa et al., 2015). In addition, it causes serious health problems in animals of agricultural interest which leads to substantial economic losses mostly in tropical and subtropical countries. Assessing the true severity of leptospirosis can be incredibly challenging, especially when early diagnosis is difficult due to nonspecific symptoms that overlap with other illnesses (Haake and Levett, 2015). Recent outbreaks of both human and canine leptospirosis in New York and California in 2020–2022 (NYC Health, 2021; Health, 2022) underscores the need for development of effective strategies to control this disease. Although serovar-specific vaccines are available for animals and at least one is available for humans, no broadly effective vaccine is available for either (Barazzone et al., 2021). The absence of an effective cross-protective vaccine candidate increases the risk of disease re-emergence on a global scale. Efforts to use leptospiral surface antigens in various vaccine formulations have shown limited success in conferring protection against leptospiral dissemination and shedding, as well as severe disease. Leptospira immune evasion strategies contribute to the complexities of finding good vaccine candidates.
The genus Leptospira is broadly categorized into two major clades P and S (Pathogens and Saprophytes) and further categorized into four subclades, P1, P2, S1, and S2 based on their virulence properties, growth conditions, and genetic make-up (Vincent et al., 2019; Picardeau, 2017). Subclade P1 is further divided in two phylogenetically related groups named P1+ (high-virulence pathogens, established pathogenic species, e.g. Leptospira interrogans) and P1- (low-virulence pathogens, phenotypically not well characterized) (Giraud-Gatineau et al., 2024). Leptospira survives in moist conditions and are free-living organisms naturally found in soil and water (Narkkul et al., 2020; Benacer et al., 2013). The spread of infection occurs through contaminated water contact with breached skin or mucosal surfaces (Ko et al., 2009). Saprophytic strains of Leptospira, such as Leptospira biflexa (S1), are unable to establish disease due to the lack of certain virulence factors (Picardeau, 2017) and have been found in natural environments around the world alongside pathogenic serovars (Vincent et al., 2019; Ko et al., 2009; Guglielmini et al., 2019). Moreover, L. biflexa exhibits certain niche-specific adaptations that allow them to persist in both environmental and host settings (Zhang et al., 2018; Castiblanco-Valencia et al., 2016).
Our previous studies (Shetty et al., 2021; Kundu et al., 2022) demonstrated that L. biflexa triggers a robust innate immune response in mice during the acute phase of infection. This raised the question of whether saprophytic Leptospira-induced immune responses could confer any degree of resistance or immune memory that could suppress a subsequent challenge with a pathogenic serovar of Leptospira. Answering that question was the main goal of the current study.
Results
Exposure to saprophytic Leptospira before infection with a pathogenic serovar prevents disease and increases survival of C3H-HeJ mice
We inoculated adult C3H-HeJ male mice with a single dose of L. biflexa 2 weeks before challenge with L. interrogans (LB1LIC1) at 8 weeks (Figure 1A) and measured a significant rescue of weight loss over a period of 15 days as compared to mice infected at 8 weeks that did not receive L. biflexa (PBS1LIC1) (p<0.0001); unchallenged control mice that received L. biflexa (LB1) or PBS (PBS1) gained weight throughout the corresponding 15 days (Figure 1B). Survival curves were generated after the mice reached the following endpoint criteria: 20% weight loss or 15 days post challenge with L. interrogans or 15 days post inoculation with L. biflexa/PBS for the controls (Figure 1C). All mice infected with L. interrogans (PBS1LIC1) reached the 20% weight loss endpoint criteria between d9 and d12 post infection. In contrast, 75% of the mice that received one dose of L. biflexa before challenge with L. interrogans (LB1LIC1) survived and gained significant body weight (Figure 1B) which was similar to the naïve control that received only PBS. Analysis of bacterial dissemination was done by qPCR of the Leptospira 16S gene in genomic DNA purified from blood, kidney tissue, and urine. Of note, although 16S rRNA primers can amplify L. biflexa 6 hr post infection (Surdel et al., 2022), we processed the tissue samples 30 days or 45 days post L. biflexa exposure. We also found that a single exposure to L. biflexa before challenge did not prevent dissemination of pathogenic L. interrogans in blood (Figure 1—figure supplement 1) or shedding in urine (Figure 1D), or kidney colonization (Figure 1E). Culture of kidney in EMJH media showed presence of ~2500 motile, morphologically intact L. interrogans under dark-field microscopy which was confirmed by 16S qPCR (Figure 1F) both on d3 and d5 post culture of kidney collected from LB1LIC1 mice; kidney from PBS1, LB1, and PBS1LIC1 did not produce positive cultures by dark-field microscopy or 16S qPCR (Figure 1—source data 1, Figure 1—figure supplement 1—source data 1).
Figure 1 with 1 supplement see all
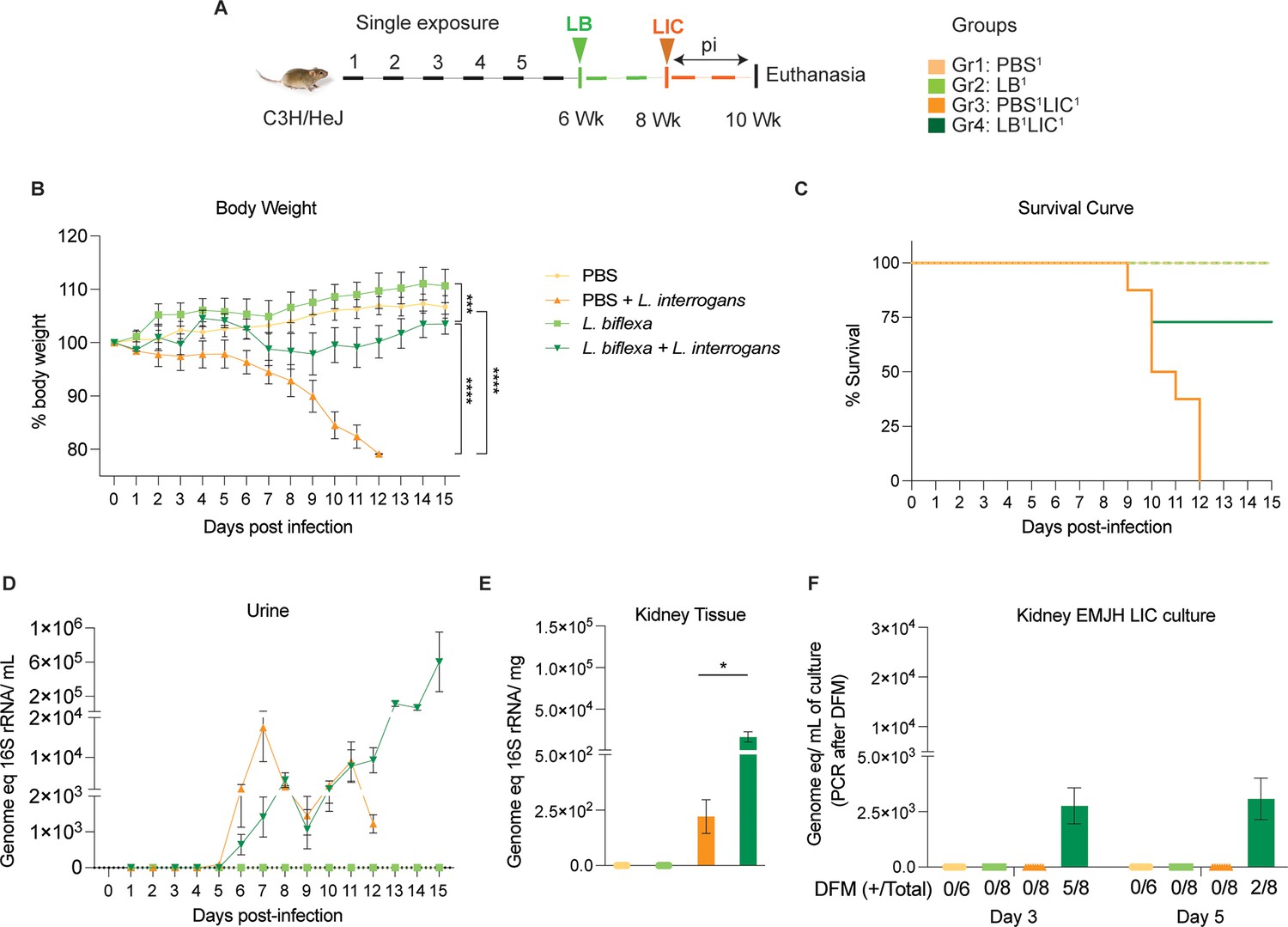
Weight loss, kidney colonization, shedding in urine, and survival to challenge with L. interrogans following a single exposure to L. biflexa.
Male C3H/HeJ mice were inoculated once with 108 L. biflexa (LB) at 6 weeks and they were challenged with 108 L. interrogans serovar Copenhageni FioCruz (LIC) at 8 weeks. (A) Experimental layout; (B) body weight measurements (%) acquired for 15 days post challenge with LIC; (C) mouse survival within the 15 days post challenge with LIC; (D) 16S rRNA qPCR quantification of live LIC in urine; (E) 16S rRNA qPCR quantification of Leptospira burden in kidney tissue harvested on d15 post challenge with LIC and (F) 16S rRNA qPCR from kidney EMJH cultures containing live Leptospira previously observed by dark-field microscopy (DFM). DFM positive culture from the total data is represented in numbers under the graph. Statistical analysis was performed by ordinary one-way ANOVA followed by Tukey’s multiple comparison correction between challenged groups and their respective controls, *p<0.05, **p<0.01, ***p<0.001, and ****p<0.0001, N=6–8 mice per group. Data represents two independent experiments.
-
Figure 1—source data 1
Excel file containing the source data used to make Figure 1.
- https://cdn.elifesciences.org/articles/96470/elife-96470-fig1-data1-v1.xlsx
In the double exposure study, mice were inoculated with two bi-weekly doses of L. biflexa 2 weeks before challenge with L. interrogans (LB2LIC2) at 10 weeks in comparison with the respective controls (Figure 2A). As expected, mice infected at 10 weeks with LIC that did not receive L. biflexa (PBS2LIC2) lost ~11% of weight on d11 post challenge and did not recover (Figure 2B). In contrast, mice that received a double dose of L. biflexa 2 weeks before challenge at 10 weeks with L. interrogans (LB2LIC2) lost a maximum of 5% weight on d10 and recovered fully by d15 post infection; unchallenged control mice that received L. biflexa (LB2) or PBS (PBS2) gained weight throughout the 15 days (Figure 2B). Survival curves generated after the mice reached endpoint criteria (Figure 2C) show that all experimental groups survived LIC infection. Analysis of bacterial dissemination showed that a double exposure to L. biflexa before challenge did not prevent dissemination of pathogenic L. interrogans in blood (Figure 2—figure supplement 1), or shedding in urine (Figure 2D), or kidney colonization (Figure 2E). Culture of kidney in EMJH media showed presence of 5000–10,000 motile, morphologically intact L. interrogans on d3 and d5 post culture of kidney collected from PBS2LIC2 mice in contrast to 1000–2500 live L. interrogans observed in culture from kidney collected from LB2LIC2 mice which was confirmed by 16S qPCR (Figure 2F); kidney from PBS2 and LB2 mice did not produce positive cultures by dark-field microscopy or 16S qPCR (Figure 2—source data 1, Figure 2—figure supplement 1—source data 1).
Figure 2 with 1 supplement see all
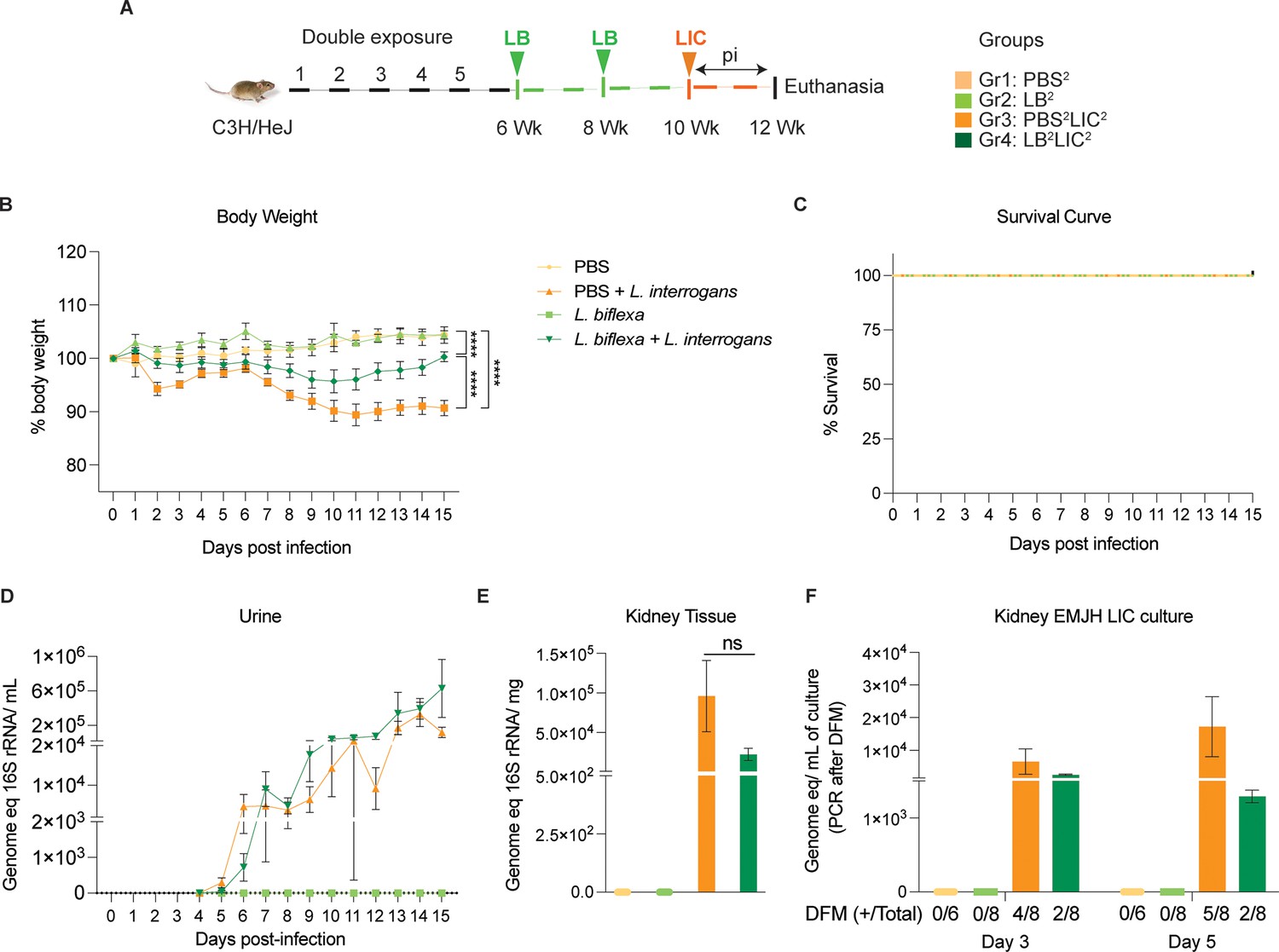
Weight loss, kidney colonization, shedding in urine, and survival to challenge with L. interrogans following a double exposure to L. biflexa.
Male C3H/HeJ mice were inoculated twice with 108 L. biflexa at 6 and 8 weeks, and at 10 weeks they were challenged with 108 L. interrogans ser Copenhageni FioCruz (LIC). (A) Experimental layout; (B) body weight measurements (%) acquired for 15 days post challenge with LIC; (C) mouse survival within the 15 days post challenge with LIC; (D) 16S rRNA qPCR quantification of live LIC in urine; (E) 16S rRNA qPCR quantification of Leptospira burden in kidney tissue harvested on d15 post challenge with LIC and (F) 16S rRNA qPCR from kidney EMJH cultures containing live Leptospira previously observed by dark-field microscopy (DFM). DFM positive culture from the total data is represented in numbers under the graph. Statistical analysis was performed by ordinary one-way ANOVA followed by Tukey’s multiple comparison correction between challenged groups and their respective controls, *p<0.05, **p<0.01, ***p<0.001, and ****p<0.0001. N=6–8 mice per group. Data represents two independent experiments.
-
Figure 2—source data 1
Excel file containing the data used to make Figure 2.
- https://cdn.elifesciences.org/articles/96470/elife-96470-fig2-data1-v1.xlsx
L. biflexa exposure before challenge with L. interrogans mitigates renal histopathological changes
As expected, hematoxylin and eosin (H&E) staining of histological slices of all kidneys from mice challenged with L. interrogans in both single and double exposure experiments (PBS1LIC1 and PBS2LIC2) showed signs of inflammation with increased immune cell infiltration (Figure 3A and C). In contrast, H&E staining of kidney slices from the groups of mice exposed to L. biflexa before L. interrogans challenge (LB1LIC1 and LB2LIC2) showed reduced immune cell infiltration. We also measured expression of a marker (ColA1) for kidney fibrosis. In both experiments, kidneys from mice challenged with L. interrogans (PBS1LIC1 and PBS2LIC2) had significantly higher expression of ColA1 as compared to the controls; in contrast, kidneys from mice challenged with L. interrogans after exposure to L. biflexa (LB1LIC1 and LB2LIC2) were not different than the controls (Figure 3B and D; Figure 3—source data 1).
Figure 3 with 1 supplement see all
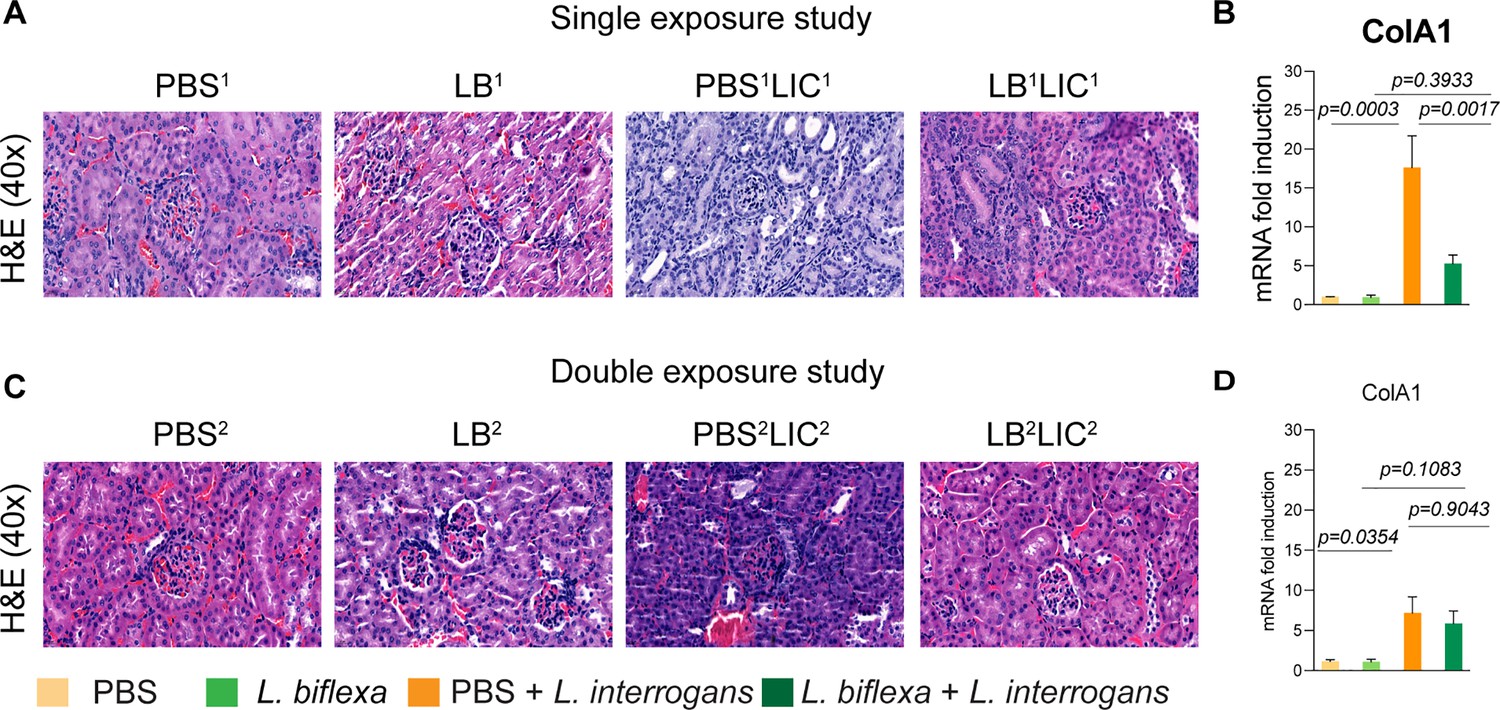
Kidney histopathology and quantification of renal fibrosis.
Representative hematoxylin and eosin (H&E)-stained kidney tissue sections from both single and double exposure studies are included in (A) and (C), respectively. The images were captured at ×40 magnification. (B) and (D) represent the mRNA expression of kidney fibrosis marker ColA1 by qPCR normalized to endogenous β-actin expression. Data was analyzed by ordinary one-way ANOVA followed by Tukey’s multiple comparison correction between challenged groups with their respective controls; *p-values are included in the graphs. Data represents one of two independent experiments.
-
Figure 3—source data 1
Excel file containing the data used to make Figure 3.
- https://cdn.elifesciences.org/articles/96470/elife-96470-fig3-data1-v1.xlsx
In addition, we were able to collect kidneys from experimental mice subjected to the double exposure of L. biflexa because they all survived subsequent challenge with L. interrogans. As such, we did a comparative gross morphological analysis between the four groups (Figure 3—figure supplement 1). We found that kidney from LB2LIC2 mice maintained their normal gross anatomy and coloration as did kidneys from control mice (PBS2 and LB2), in contrast to kidneys from mice challenged with L. interrogans that were not previously exposed to L. biflexa (PBS2LIC2).
Serologic IgG2a responses to L. interrogans were significantly higher in mice pre-exposed to L. biflexa before challenge with L. interrogans
In both single and double L. biflexa exposure experiments we measured anti-L. interrogans total IgM, total IgG, and IgG subtypes IgG1, IgG2a, and IgG3 in serum collected 2 weeks after challenge with L. interrogans (IgG subtypes shown in Figure 4A and B). In both experiments, total IgM and IgG were significantly increased in PBS-LIC and LB-LIC when compared to the respective controls, but not between PBS-LIC and LB-LIC groups. Regarding IgG isotypes, IgG1 was generally low and IgG2a as well as IgG3 were generally high in groups infected with L. interrogans. Although differences between groups (PBS2v PBS2LIC2 and LB1/2 vs LB1/2LIC1/2) were significant, differences between the LIC infected groups (PBS1/2LIC1/2 vs LB1/2LIC1/2) were not significant for IgG1 and for IgG3 in contrast to IgG2a (p=0.001 for single exposure and p=0.0095 for double exposure) (Figure 4—source data 1).
Figure 4
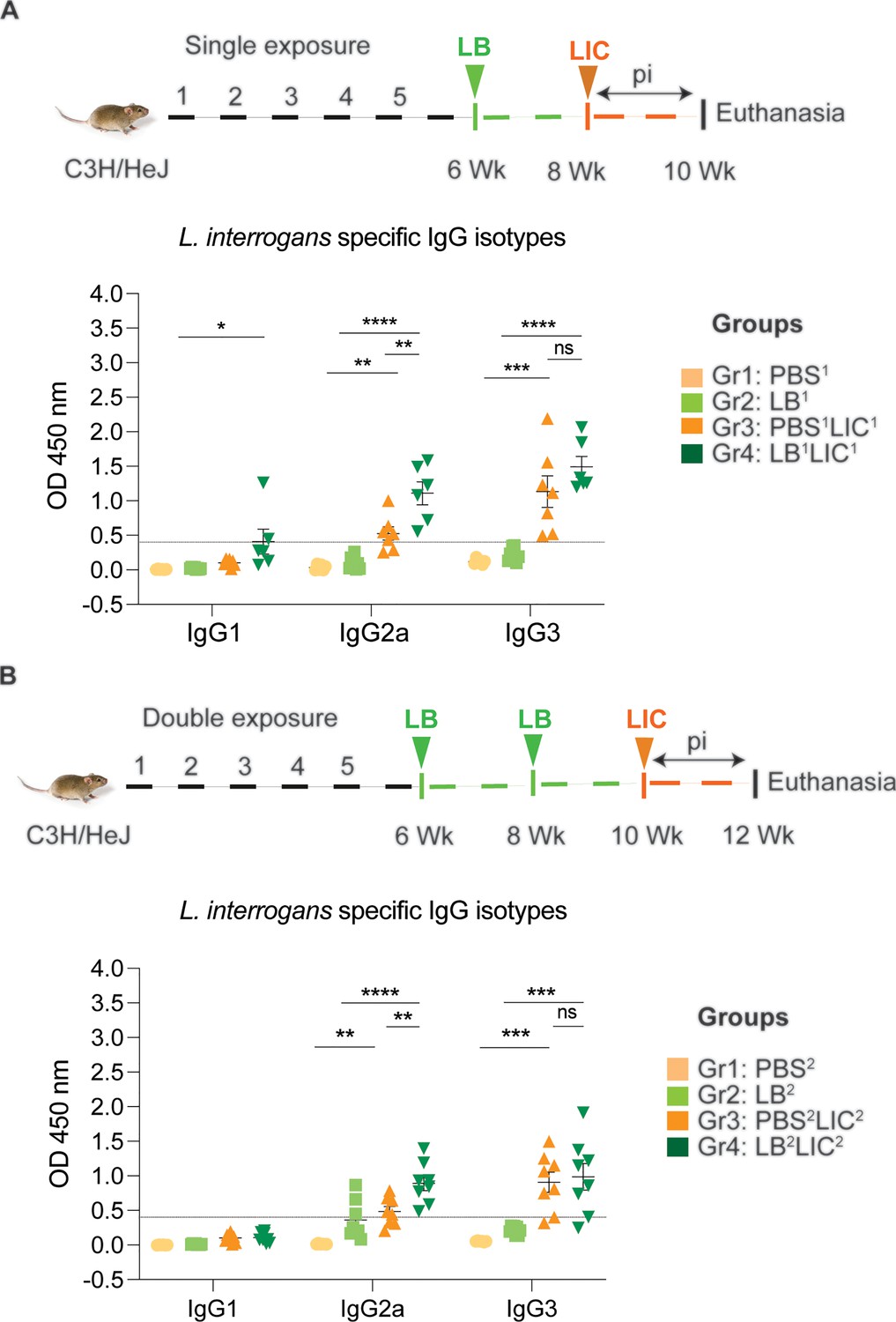
Detection of IgG1, IgG2a, and IgG3 specific to L. interrogans in serum from experimental mice.
(A) represents IgG isotypes specific to L. interrogans in 10-week serum of mice exposed once to L. biflexa before L. interrogans challenge. (B) represents IgG isotypes specific to L. interrogans in 12-week serum of mice exposed twice to L. biflexa before L. interrogans challenge. Ordinary one-way ANOVA followed by Tukey’s multiple comparison correction test was used to compare between challenged groups with their respective controls; *p<0.05, **p<0.01, ***p<0.001, ****p<0.0001, and ns = not significant; N=6–8 mice per group. Data represents two independent experiments.
-
Figure 4—source data 1
Excel file containing the raw data points used to make Figure 4.
- https://cdn.elifesciences.org/articles/96470/elife-96470-fig4-data1-v1.xlsx
Exposure to non-pathogenic L. biflexa before pathogenic L. interrogans challenge induced increased frequencies of effector helper T cells in spleen
We immunophenotyped the spleen cells from the mice subjected to double L. biflexa exposure because all animals survived to the term of the experiment. The gating strategy used for spleen cell immunophenotyping is provided in Figure 5—figure supplement 1. Mice in the single exposure experiment met endpoint criteria before the term of the experiment and thus we were not able to process spleen for immunophenotyping. In the L. biflexa double exposure experiment, we measured increased frequencies in B cells when LIC infected mice were compared to the respective controls, but not between PBS2LIC2 and LB2LIC2 mice (Figure 5A). We measured decreased frequencies in T cells when LIC infected mice were compared to the respective controls but not between PBS2LIC2 and LB2LIC2 mice (Figure 5B). No differences were observed in NK cells between any of the groups (Figure 5C). We measured increased frequencies in helper T cells between all groups; of note, PBS2LIC2 vs LB2LIC2 p=0.006 (Figure 5D). We also measured decreased frequencies in cytotoxic T cells between all groups; of note PBS2LIC2 vs LB2LIC2 p=0.0056 (Figure 5E; Figure 5—source data 1).
Figure 5 with 1 supplement see all
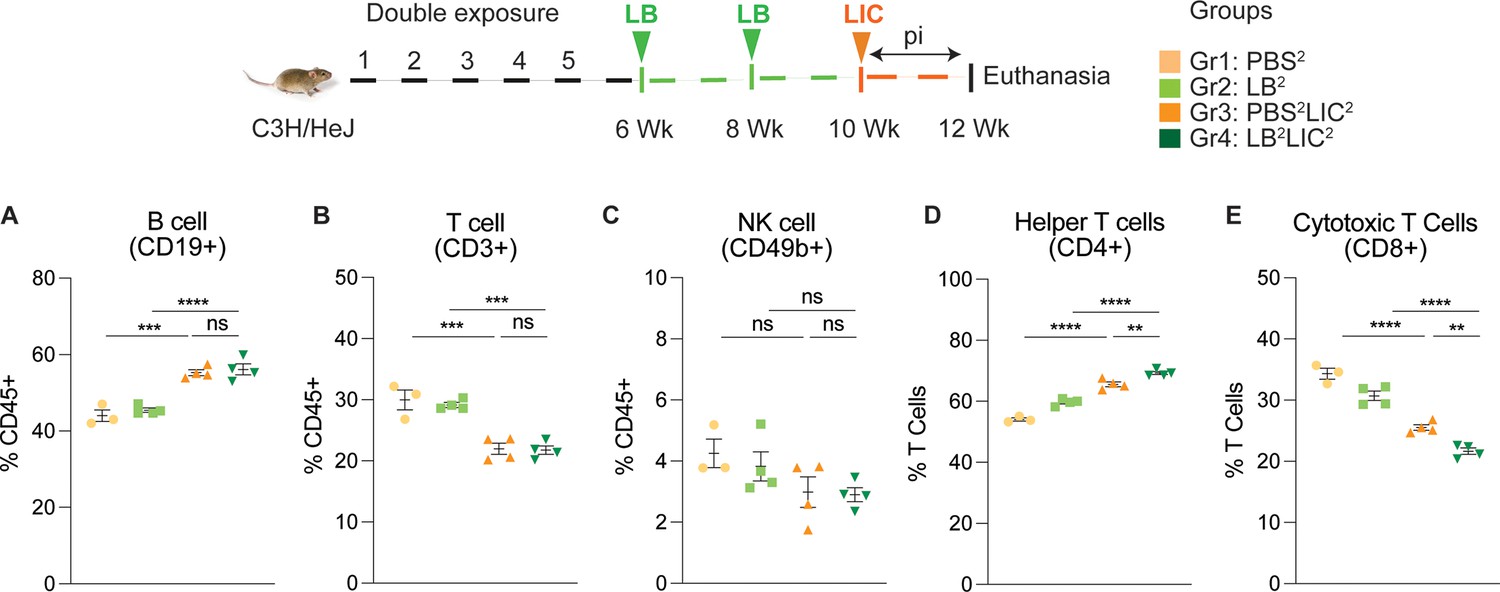
Frequency of lymphocytes in spleen of mice subjected to a double exposure of L. biflexa before challenge with L. interrogans.
(A–E) show B cell (CD19+), T cell (CD3+), NK cell (CD49b+), helper T cell (CD4+), and cytotoxic T cell (CD8+) frequencies in groups of experimental mice. Ordinary one-way ANOVA followed by Tukey’s multiple comparison correction test was used to compare between challenged groups and their respective controls; **p<0.01, ***p<0.001, ****p<0.0001, and ns = not significant; N=3–4 mice per group. Data represents one of two independent experiments.
-
Figure 5—source data 1
Excel file containing data points used to generate Figure 5.
- https://cdn.elifesciences.org/articles/96470/elife-96470-fig5-data1-v1.xlsx
Furthermore, T cell subset typing (Figure 6) showed that frequency of early effector CD4+ T helper cells (Figure 6B, CD44-CD62L-) and effector T helper cells (Figure 6C, CD44+CD62L-) were significantly increased when compared between the LIC challenged groups and the respective controls (except PBS2 vs PBS2LIC2 early effectors) and that frequency of early effector and effector T helper cells was higher in the LB2LIC2 group than PBS2LIC2. No major changes were measured in memory CD4+ T helper cells (Figure 6D, CD44+CD62L+). In the CD8+ cytotoxic T cell subsets, we measured significant decreases in frequency of naïve T cells between LIC infected groups and the respective controls (Figure 6E, CD62L+CD44-) and this was replicated in the CD8+ cytotoxic memory except that differences with the LIC infected groups were not significant (Figure 6H, CD44+CD62L+) (Figure 6—source data 1).
Figure 6
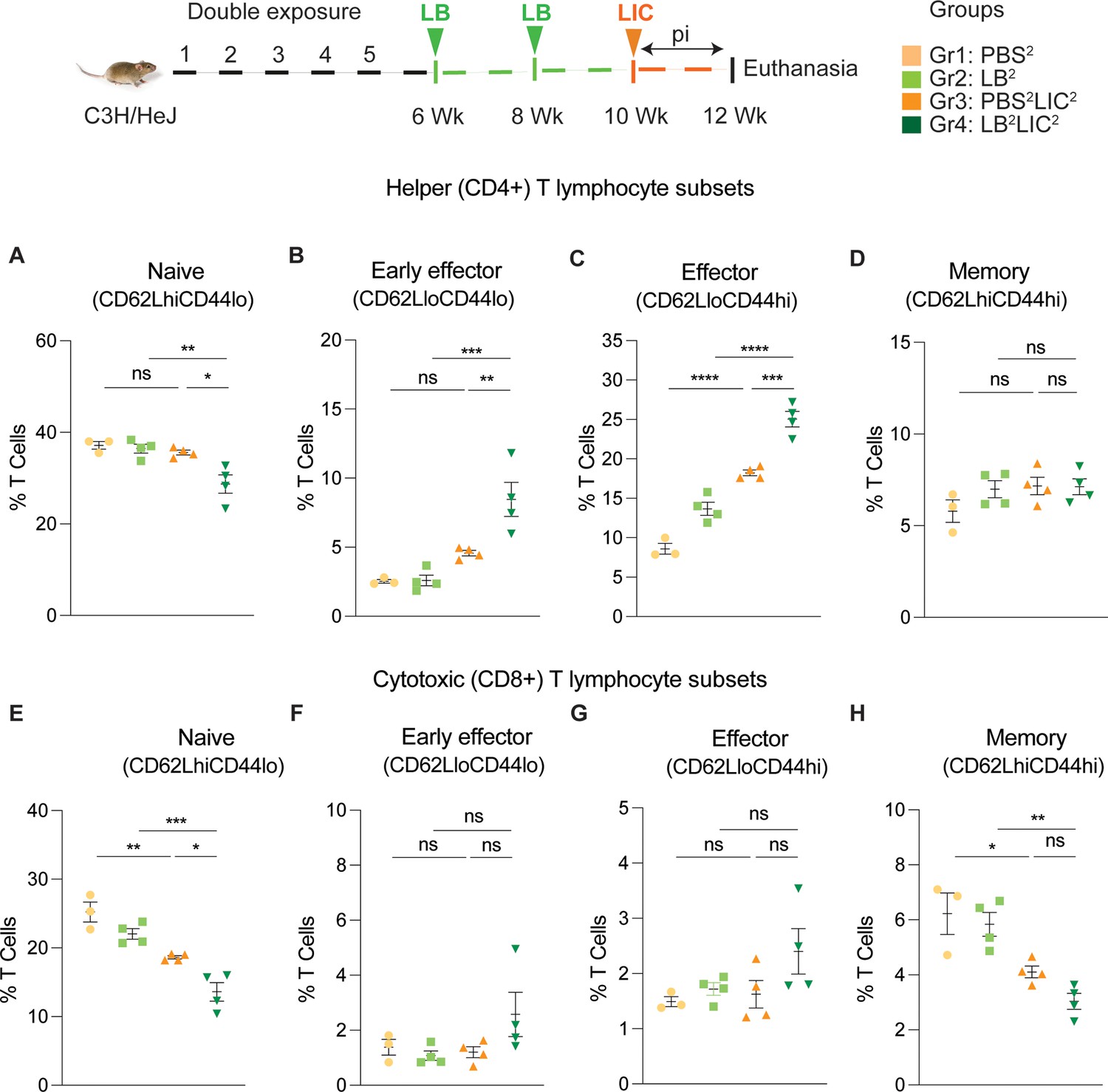
Frequency of T cell subsets (CD62L/CD44) in spleen of mice subjected to a double exposure of L. biflexa before challenge with L. interrogans.
(A–D) represent naïve, early effector, effector, and memory subsets of CD4+ helper T lymphocytes, respectively. (E–H) represent naive, early effector, effector, and memory subsets of CD8+ cytotoxic T lymphocytes, respectively. Ordinary one-way ANOVA followed by Tukey’s multiple comparison correction test was used to compare between challenged groups and their respective controls; *p<0.05, **p<0.01, ***p<0.001, ****p<0.0001, and ns = not significant; N=3–4 mice per group. Data represents one of two independent experiments.
-
Figure 6—source data 1
Excel file containing the source data used to generate Figure 6.
- https://cdn.elifesciences.org/articles/96470/elife-96470-fig6-data1-v1.xlsx
Discussion
Understanding host immune responses to Leptospira infection is crucial for advancing our ability to develop new control measures for leptospirosis (Wunder et al., 2021; Vernel-Pauillac et al., 2021; Potula et al., 2017; Fortes-Gabriel et al., 2022; Santecchia et al., 2019; Murray et al., 2018). Given their widespread presence in nature, the likelihood of humans or animals getting exposed to non-pathogenic serovars of Leptospira is likely high (Benacer et al., 2013; Ko et al., 2009). Our previous studies showed innate immunity engagement during saprophytic L. biflexa infection in mice (Shetty et al., 2021; Kundu et al., 2022). In addition, L. biflexa extracts can be used to detect Leptospira-specific antibody in up to 67% of serum from patients with clinically confirmed leptospirosis (Fortes-Gabriel et al., 2022) which points to a high degree of immunodominant cross-reactive epitopes between L. biflexa and pathogenic Leptospira. Further, the current hypothesis regarding evolution of Leptospira species is that symbiosis of Leptospira with eukaryotes emerged from free-living ancestral species (Thibeaux et al., 2018); in other words, pathogenic Leptospira may have evolved from an environmental ancestor (Vincent et al., 2019). Thus, we hypothesized that these highly cross-reactive immunodominant epitopes may also induce cross-protective immune responses. The objective of the current study was to assess whether exposure to a live saprophytic serovar of Leptospira provides any heterologous cross-species protection against a subsequent challenge with a pathogenic serovar in a mammalian host (mouse).
In the initial analysis of pathogenesis (Figure 1, Figure 2) we observed that prior exposure to one or two doses of saprophytic L. biflexa rescues weight loss in mice challenged with pathogenic L. interrogans at 8 weeks (LB1LIC1) and at 10 weeks (LB2LIC2), respectively. Weight gain correlated with survival (75% survival in LB1LIC1 vs 0% survival of the PBS1LIC1 group) in the single exposure experiment, where we expected all mice infected at 8 weeks with pathogenic L. interrogans (PBS1LIC1) to irreversibly lose weight and meet endpoint criteria for euthanasia before the 2-week term of the experiment (Fortes-Gabriel et al., 2022, Shetty et al., 2022). Loss of mice due to irreversible weight loss is not expected if mice are infected with LIC at 10 weeks of age (Nair et al., 2020), as observed in the L. interrogans control group in the double exposure experiment (PBS2LIC2). In both experiments, mice exposed to L. biflexa before challenge with LIC produced evidence of L. interrogans dissemination in blood, shedding in urine, and kidney colonization.
Histological inspection of kidney slices (Figure 3) showed that exposure to a saprophytic Leptospira before challenge supported normal structural morphology and prevented infiltration of immune cells in both single and double exposure experiments; in addition, it significantly reduced a fibrosis marker (ColA1) in the single exposure experiment. In the double exposure experiment, differences in ColA1 fibrosis marker are not significant between the two LIC infected groups because 10-week-old C3H-HeJ infected with LIC are more resistant to pathology resultant from infection. Our findings are intriguing as they suggest that while prior live saprophytic exposure did not prevent infection or leptospiral dissemination, it may confer protection against kidney fibrosis.
Our data also shows that prior exposure to non-pathogenic Leptospira before pathogenic challenge induced higher antibody titer in the serum, specifically IgG2a antibodies against L. interrogans in both single and double exposure experiments (Figure 4). Increased IgG2a response in serum is associated with induction of a Th1 biased immune response. Others have recently found that saprophytic L. biflexa induced Th1 responses, higher T cell proliferation, and IFN-γ producing CD4+ T cells (Krangvichian et al., 2023). Persistent IgM and strain-specific IgG responses were observed during a homologous leptospiral challenge in C57BL6/J mice (Vernel-Pauillac et al., 2021). In our study, exposure to a saprophytic Leptospira induced antibody responses that may provide heterologous protection against the pathogenic strain of Leptospira. This supports a promising broad-spectrum efficacy. Thus, live vaccines derived from a saprophytic strain of Leptospira could offer broader protection and overcome the limitation of serovar specificity often observed with killed whole-cell vaccines based on pathogenic strains.
Differences in antibody titer among the L. interrogans infected group pre-exposed to saprophytic L. biflexa can be attributed to the robust trafficking and differentiation of B and helper T cells (CD4+) measured in spleen (Figure 5 and Figure 6). Presence of effector helper T (CD4+) cells in the spleen indicate a robust cellular immune response as these cells produce cytokines that play a pivotal role in activating other immune cells, including antibody-producing B cells. Moreover, our findings align with another observation which further reinforces the potential immunostimulatory properties of components (polar lipids) derived from saprophytic L. biflexa, indicating that these components could play a crucial role in inducing robust B cell responses (Faisal et al., 2009). Induction of helper T cell responses along with dynamic transition from naïve to early effector and effector without T helper memory reflects an orchestrated immune response upon pathogenic challenge in the saprophytic pre-exposed group that is typical of effective responses to vaccines. Previous studies have highlighted the significance of activated CD4+ T cells during Leptospira infection in providing protective immunity to the host and mitigating the severity of leptospirosis by releasing cytokines (Volz et al., 2015). Correlating induction of chemo-cytokines by saprophytic Leptospira with subsequent adaptive immune responses, such as the activation of CD4+ T cells or the production of specific antibodies, provides insights into how innate immune signals drive the adaptive immune response against a pathogenic threat. It may also aid in identifying key signaling molecules or pathways that could be targeted for therapeutic interventions or vaccine design.
While other researchers have explored vaccination strategies using live attenuated or mutant strains of pathogenic serovars, our approach was to utilize a live saprophytic bacterial strain which is unique in the field (Wunder et al., 2021; Potula et al., 2017; Lauretti-Ferreira et al., 2020; Teixeira et al., 2019). We previously showed that oral delivery of a probiotic strain, Lactobacillus plantarum, reduces the severity of leptospirosis by recruiting myeloid cells (Fortes-Gabriel et al., 2022) which suggests that a general phenomenon of trained immunity may be involved. Current vaccines based on inactivated pathogenic species provide equivalent protection to the one achieved in this study (Barazzone et al., 2021; Vernel-Pauillac and Werts, 2018) with the caveat of being serovar-specific (Teixeira et al., 2019; Vernel-Pauillac and Werts, 2018; Adler, 2015). Although our current study conclusively shows protection from severe leptospirosis after heterologous challenge, it remains to be shown if protection extends to multiple pathogenic serovars of Leptospira. Using a live saprophytic strain of Leptospira as control strategy could pave the way for development of novel broadly effective vaccines against leptospirosis. Such a vaccine could have a substantial economic impact if applied to animals of agricultural interest.
Another interesting aspect of our current study is that it shows that exposure to a live saprophytic strain of Leptospira provides protection against a pathogenic serovar. Thus, in the real-life scenario where individuals or animals may naturally encounter a saprophytic Leptospira species, they may develop immune responses that mitigate severe disease outcomes if the host later encounters a pathogenic strain of Leptospira. By exploring the immune dynamics during the co-exposure to different Leptospira serovars, this study could open avenues of research on strategies that leverage natural exposure to saprophytic species to devise safe control measures against leptospirosis. This concept is important for understanding the epidemiological risk factors of leptospirosis and it should be applicable to other infectious diseases caused by direct contact between the pathogen and mucosal membranes or abraded host skin.
Importantly, we found that in mice pre-exposed to live saprophytic Leptospira, there was a correlation between kidney health after LIC infection (less infiltration of immune cells in kidney and less fibrosis marker ColA1) and higher shedding of live LIC in urine. This suggest that a status of homeostasis was reached after kidney colonization that helps the spirochete complete its enzootic cycle. Additional research is needed to fully understand the mechanisms involved in kidney homeostasis after LIC infection.
Materials and methods
Animals
Male C3H/HeJ mice (n=6–8/group) were purchased from The Jackson Laboratory (Bar Harbor, ME, USA) and were maintained in a pathogen-free environment in the Laboratory Animal Care Unit of the University of Tennessee Health Science Center (UTHSC). All experiments were performed in compliance with the UTHSC Institutional Animal Care and Use Committee (IACUC), Protocol no. 19-0062.
Bacteria
Non-pathogenic L. biflexa serovar Patoc (LB) belonging to subclade S1 was purchased from ATCC and grown in EMJH media. Pathogenic L. interrogans serovar Copenhageni strain Fiocruz L1-130 (LIC) belonging to subclade P1+ (high-virulence pathogens) (Vincent et al., 2019; Giraud-Gatineau et al., 2024) was grown in EMJH media and subsequently passaged in hamster to maintain virulence. EMJH culture passage 2 was used to inoculate mice (108) after counting Leptospira under a dark-field microscope (Zeiss USA, Hawthorne, NY, USA) using a Petroff-Hausser chamber.
Infection of mice and study design
Request a detailed protocolWe carried out two experiments set apart by a single or double exposure to a saprophytic serovar of Leptospira (L. biflexa) before challenge with a pathogenic serovar (L. interrogans). Groups of mice were inoculated with 108 Leptospira intraperitoneally (IP) both for exposure to L. biflexa and for challenge with L. interrogans. Each experiment was reproduced once. In the single exposure study (Figure 1A), Group 1 (n=3) was the naive control which received PBS (PBS1), Group 2 (n=4) was inoculated with 108 L. biflexa once at 6 weeks (LB1), Group 3 (n=4) received PBS for 2 weeks followed by challenge with 108 L. interrogans at 8 weeks (PBS1LIC1), and Group 4 (n=4) was inoculated with 108 L. biflexa at 6 weeks and challenged with 108 L. interrogans at 8 weeks (LB1LIC1). In the double exposure study (Figure 2A), Group 1 (n=3) was the naive control which received PBS (PBS2), Group 2 (n=4) received 108 L. biflexa IP at 6 and 8 weeks (LB2), Group 3 (n=4) received PBS for 2 weeks followed by challenge with 108 L. interrogans at 10 weeks (PBS2LIC2), and Group 4 (n=4) was inoculated with 108 L. biflexa at 6 and 8 weeks and challenged with 108 L. interrogans at 10 weeks (LB2LIC2). Weight was monitored daily. Mice were euthanized 15 days after L. interrogans challenge or when they reached the endpoint criteria (20% body weight loss post infection). Blood and kidney were collected at euthanasia: blood was used for quantification of anti-Leptospira antibody; kidney was used for quantification of Leptospira load (16S rRNA) and it was cultured in EMJH media for evaluation of bacterial viability. Furthermore, kidney samples were stored in 10% formalin for H&E staining. Spleen for flow cytometric analysis was collected from mice after euthanasia only in the double exposure study given that all mice consistently survived challenge.
Leptospira detection through qPCR
Request a detailed protocolIsolation of genomic DNA from blood, urine, and kidney were carried out using NucleoSpin tissue kit (Clontech, Mountain View, CA, USA) according to the manufacturer’s instructions. Leptospira 16S rRNA primers (Forward- CCCGCGTCCGATTAG and Reverse- TCCATTGTGGCCGAACAC) and TAMRA probe (CTCACCAAGGCGACGATCGGTAGC) were used for detection of Leptospira genus using qPCR with a standard curve of 105 to 1 L. interrogans (Nair et al., 2020, Richer et al., 2015). Similarly, qPCR was performed with kidney tissues placed in EMJH to grow live Leptospira after culturing for 5 days and visually quantified under a dark-field microscope (20×, Zeiss USA, Hawthorne) on d3 and d5 post culture inoculation.
RNA isolation and RT-PCR
Request a detailed protocolKidneys were stored in RNA later after euthanasia. RNeasy Mini Kit (QIAGEN) was used to extract total RNA from kidney tissue according to the manufacturer’s specifications. RNA purity was measured using a Nanodrop instrument (Thermo Scientific) at A260/280 ratio. cDNA was prepared using cDNA reverse transcription kit (Applied Biosystems). ColA1 primers (Forward- TAAGGGTACCGCTGGAGAAC, Reverse- GTTCACCTCTCTCACCAGCA), TAMRA probe (AGAGCGAGGCCTTCCCGGAC), and β-actin primers (Forward- CCACAGCTGAGAGGGAAATC, Reverse- CCAATAGTGATGACCTGGCCG), TAMRA probe (GGAGATGGCCACTGCCGCATC) were purchased from Eurofins Genomics.
Histopathology by H&E staining
Request a detailed protocolKidney tissues were fixed in formalin buffer. Histopathology was performed at the Histology Department, UT Methodist University Hospital, Memphis, TN. Digital scanning of inflammatory cell infiltration was measured by taking images of ~5 fields per sample under ×20 magnification. Images were captured after digitally scanning the H&E slides using Panoramic 350 Flash III (3D Histech, Hungary) and CaseViewer software.
ELISA
Request a detailed protocolLeptospiral extract for L. biflexa and L. interrogans were prepared as described previously (Fortes-Gabriel et al., 2022). Briefly, Leptospira was cultured in EMJH media and once confluency was observed, cells were centrifuged to obtain a pellet. This pellet was then incubated with BugBuster solution (1 mL) at room temperature (RT) in a shaker incubator (100 rpm) for 20 min and homogenized by vortexing. Stocks were stored at –20°C. This whole-cell extract of Leptospira was then diluted in 1× sodium carbonate coating buffer. Nunc MaxiSorp flat-bottom 96-well plates (eBioscience, San Diego, CA, USA) were coated with extracts prepared from 107 to 108 bacteria per well and incubated at 4°C overnight. Cells were washed using 1× PBST the following day and blocked for 1 hr using 1% BSA solution, followed by another wash with 1× PBST. Serum samples (1:100) were added to the antigen-coated wells and incubated at 37°C for 1 hr, washed twice with 1× PBST, followed by HRP conjugate secondary anti-mouse- IgG1, IgG2a, and IgG3 (1:10,000) which was incubated for 30 min. After washing the plate three times with 1× PBST the color was developed using TMB SureBlue followed by Stop solution before the absorbance was measured at OD 450 nm using an ELISA plate reader (Molecular Devices Spetramax).
Flow cytometry
Request a detailed protocolSpleens were chopped into small pieces and macerated to prepare single-cell suspensions on the same day of euthanasia to avoid loss of cell viability. RBC lysis was performed using ACK lysis buffer (Gibco). AO/PI dual staining was used to count live/dead cells on a Luna counter (Logos Biosystems, South Korea). 106 cells were seeded in a 96-well microtiter plate after washing with 1× PBS twice. Blocking was performed with anti-mouse CD16/32 antibody (1:100), followed by 20 min incubation on ice. Fluorochrome-conjugated antibodies (Supplementary file 1) were used to stain specific cell surface markers after 30 min incubation in the dark at 4°C. Freshly prepared flow staining buffer was used for washing stained cells. Cells were fixed using 4% paraformaldehyde for 10–15 min at room temperature. Beads were stained using specific fluorochrome-conjugated antibodies and used for compensation, while FMO prepared with spleen cells simultaneously were used for gating controls. Cells were resuspended in flow staining buffer and the Bio-Rad ZE5 cell analyzer was used for data acquisition. Data analysis was done using FlowJo software.
Statistical analysis
Request a detailed protocolOne-way ANOVA with Tukey’s multiple comparison test and unpaired t-test with Welch’s correction were used to analyze differences between experimental groups. GraphPad Prism software was used to plot graphs; a value of p<0.05 is considered significant. p-Values from all figures for the different experimental groups analyzed by one-way ANOVA and compared with Tukey’s multiple comparison test are provided in Supplementary file 2.
Data availability
All data generated or analyzed during this study are included in this manuscript and source data files.
References
-
Vaccines against leptospirosisCurrent Topics in Microbiology and Immunology 387:251–272.https://doi.org/10.1007/978-3-662-45059-8_10
-
Pathogenic and saprophytic Leptospira species in water and soils from selected urban sites in peninsular MalaysiaMicrobes and Environments 28:135–140.
-
Global morbidity and mortality of leptospirosis: A systematic reviewPLOS Neglected Tropical Diseases 9:e0003898.https://doi.org/10.1371/journal.pntd.0003898
-
Genus-wide Leptospira core genome multilocus sequence typing for strain taxonomy and global surveillancePLOS Neglected Tropical Diseases 13:e0007374.https://doi.org/10.1371/journal.pntd.0007374
-
Leptospirosis in humansCurrent Topics in Microbiology and Immunology 387:65–97.https://doi.org/10.1007/978-3-662-45059-8_5
-
Leptospira: the dawn of the molecular genetics era for an emerging zoonotic pathogenNature Reviews. Microbiology 7:736–747.https://doi.org/10.1038/nrmicro2208
-
Impaired functions of human monocyte-derived dendritic cells and induction of regulatory T cells by pathogenic LeptospiraPLOS Neglected Tropical Diseases 17:e0011781.https://doi.org/10.1371/journal.pntd.0011781
-
Necroptosis contributes to persistent inflammation during acute leptospirosisFrontiers in Immunology 13:810834.https://doi.org/10.3389/fimmu.2022.810834
-
Heterologous protection elicited by a live, attenuated, Leptospira vaccineVeterinary Microbiology 223:47–50.https://doi.org/10.1016/j.vetmic.2018.07.018
-
Virulence of the zoonotic agent of leptospirosis: still terra incognita?Nature Reviews. Microbiology 15:297–307.https://doi.org/10.1038/nrmicro.2017.5
-
Mouse model for sublethal Leptospira interrogans infectionInfection and Immunity 83:4693–4700.https://doi.org/10.1128/IAI.01115-15
-
Inflammatory Signatures of Pathogenic and Non-Pathogenic Leptospira Infection in Susceptible C3H-HeJ MiceFrontiers in Cellular and Infection Microbiology 11:677999.https://doi.org/10.3389/fcimb.2021.677999
-
Transient presence of live leptospira interrogans in murine testesMicrobiology Spectrum 10:e0277521.https://doi.org/10.1128/spectrum.02775-21
-
Hematogenous dissemination of pathogenic and non-pathogenic Leptospira in a short-term murine model of infectionFrontiers in Cellular and Infection Microbiology 12:917962.https://doi.org/10.3389/fcimb.2022.917962
-
Revisiting the taxonomy and evolution of pathogenicity of the genus Leptospira through the prism of genomicsPLOS Neglected Tropical Diseases 13:e0007270.https://doi.org/10.1371/journal.pntd.0007270
-
Specific CD4+ T-cell reactivity and cytokine release in different clinical presentations of leptospirosisClinical and Vaccine Immunology 22:1276–1284.https://doi.org/10.1128/CVI.00397-15
Article and author information
Author details
Maria Gomes-Solecki

Funding
National Institute of Allergy and Infectious Diseases (R01 AI139267)
- Suman Kundu
- Maria Gomes-Solecki
National Institute of Allergy and Infectious Diseases (R21 AI142129)
- Suman Kundu
- Advait Shetty
- Maria Gomes-Solecki
The funders had no role in study design, data collection and interpretation, or the decision to submit the work for publication.
Acknowledgements
We sincerely thank Dr. Diedre Daria and Dr. Tony Marion of the Flow Cytometry and Cell Sorting Core facility at UTHSC. We would like to acknowledge the Histology department from UT Methodist University Hospital for processing and staining tissues, Michelle Morrison from the Department of Pathology, UTHSC for digitally scanning tissue slides for histology analysis. This work was supported by the National Institute of Allergy and Infectious Diseases (NIAID), United States National Institutes of Health (NIH), grant numbers R01 AI139267 (MGS), R21 AI 142129 (MGS). The content of this manuscript is totally the responsibility of the authors and does not involve the official views of NIAID or NIH.
Ethics
This study was performed in strict accordance with the recommendations in the Guide for the Care and Use of Laboratory Animals of the National Institutes of Health. All of the animals were handled according to approved institutional animal care and use committee (IACUC) protocol (#19-0062) of the University of Tennessee Health Science Center (UTHSC).
Version history
- Sent for peer review:
- Preprint posted:
- Reviewed Preprint version 1:
- Reviewed Preprint version 2:
- Version of Record published:
Cite all versions
You can cite all versions using the DOI https://doi.org/10.7554/eLife.96470. This DOI represents all versions, and will always resolve to the latest one.
Copyright
© 2024, Kundu, Shetty et al.
This article is distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use and redistribution provided that the original author and source are credited.
Metrics
-
- 464
- views
-
- 40
- downloads
-
- 2
- citations
Views, downloads and citations are aggregated across all versions of this paper published by eLife.
Download links
A two-part list of links to download the article, or parts of the article, in various formats.
Downloads (link to download the article as PDF)
Open citations (links to open the citations from this article in various online reference manager services)
Cite this article (links to download the citations from this article in formats compatible with various reference manager tools)
Exposure to live saprophytic Leptospira before challenge with a pathogenic serovar prevents severe leptospirosis and promotes kidney homeostasis
eLife 13:RP96470.
https://doi.org/10.7554/eLife.96470.3
Further reading
-
- Cell Biology
- Immunology and Inflammation
Macrophages are crucial in the body’s inflammatory response, with tightly regulated functions for optimal immune system performance. Our study reveals that the RAS–p110α signalling pathway, known for its involvement in various biological processes and tumourigenesis, regulates two vital aspects of the inflammatory response in macrophages: the initial monocyte movement and later-stage lysosomal function. Disrupting this pathway, either in a mouse model or through drug intervention, hampers the inflammatory response, leading to delayed resolution and the development of more severe acute inflammatory reactions in live models. This discovery uncovers a previously unknown role of the p110α isoform in immune regulation within macrophages, offering insight into the complex mechanisms governing their function during inflammation and opening new avenues for modulating inflammatory responses.
-
- Immunology and Inflammation
- Microbiology and Infectious Disease
Pseudomonas aeruginosa (PA) is an opportunistic, frequently multidrug-resistant pathogen that can cause severe infections in hospitalized patients. Antibodies against the PA virulence factor, PcrV, protect from death and disease in a variety of animal models. However, clinical trials of PcrV-binding antibody-based products have thus far failed to demonstrate benefit. Prior candidates were derivations of antibodies identified using protein-immunized animal systems and required extensive engineering to optimize binding and/or reduce immunogenicity. Of note, PA infections are common in people with cystic fibrosis (pwCF), who are generally believed to mount normal adaptive immune responses. Here, we utilized a tetramer reagent to detect and isolate PcrV-specific B cells in pwCF and, via single-cell sorting and paired-chain sequencing, identified the B cell receptor (BCR) variable region sequences that confer PcrV-specificity. We derived multiple high affinity anti-PcrV monoclonal antibodies (mAbs) from PcrV-specific B cells across three donors, including mAbs that exhibit potent anti-PA activity in a murine pneumonia model. This robust strategy for mAb discovery expands what is known about PA-specific B cells in pwCF and yields novel mAbs with potential for future clinical use.




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}