A two-lane mechanism for selective biological ammonium transport
- Strathclyde Institute of Pharmacy and Biomedical Sciences, University of Strathclyde, United Kingdom
- Computational Biology, School of Life Sciences, University of Dundee, United Kingdom
- Physics, School of Science and Engineering, University of Dundee, United Kingdom
- Biology of Membrane Transport Laboratory, Department of Molecular Biology, Université Libre de Bruxelles, Belgium
Abstract
The transport of charged molecules across biological membranes faces the dual problem of accommodating charges in a highly hydrophobic environment while maintaining selective substrate translocation. This has been the subject of a particular controversy for the exchange of ammonium across cellular membranes, an essential process in all domains of life. Ammonium transport is mediated by the ubiquitous Amt/Mep/Rh transporters that includes the human Rhesus factors. Here, using a combination of electrophysiology, yeast functional complementation and extended molecular dynamics simulations, we reveal a unique two-lane pathway for electrogenic NH4+ transport in two archetypal members of the family, the transporters AmtB from Escherichia coli and Rh50 from Nitrosomonas europaea. The pathway underpins a mechanism by which charged H+ and neutral NH3 are carried separately across the membrane after NH4+ deprotonation. This mechanism defines a new principle of achieving transport selectivity against competing ions in a biological transport process.
Introduction
The transport of ammonium across cell membranes is a fundamental biological process in all domains of life. Ammonium exchange is mediated by the ubiquitous ammonium transporter/methylammonium-ammonium permease/Rhesus (Amt/Mep/Rh) protein family. The major role of bacterial, fungal, and plant Amt/Mep proteins is to scavenge ammonium for biosynthetic assimilation, whereas mammals are thought to produce Rh proteins in erythrocytes, kidney, and liver cells for detoxification purposes and to maintain pH homeostasis (Biver et al., 2008; Huang and Ye, 2010). In humans, Rh mutations are linked to pathologies that include inherited hemolytic anemia, stomatocytosis, and early-onset depressive disorder (Huang and Ye, 2010). Despite this key general and biomedical importance, so far, no consensus on the pathway and mechanism of biological ammonium transport has been reached.
High-resolution structures available for several Amt, Mep and Rh proteins show a strongly hydrophobic pore leading towards the cytoplasm (Andrade et al., 2005; Gruswitz et al., 2010; Khademi et al., 2004; Lupo et al., 2007; van den Berg et al., 2016). This observation led to the conclusion that the species translocated through Amt/Mep/Rh proteins is neutral NH3. However, this view has been experimentally challenged, first for some plant Amt proteins (Ludewig et al., 2002; Mayer et al., 2006; McDonald and Ward, 2016; Neuhäuser et al., 2014), followed by further in-vitro studies revealing that the activity of bacterial Amt proteins is electrogenic (Mirandela et al., 2019; Wacker et al., 2014). Taken together, these findings renewed a long-standing debate on the mechanism by which a charged molecule is translocated through a hydrophobic pore and how selectivity for NH4+ over competing ions is achieved.
Here, we reveal the pathways, mechanism, and key determinants of selectivity of electrogenic ammonium transport in Amt and Rh proteins, unifying the diverse observations that led to these seemingly incompatible suggestions. The transport mechanism is underpinned by the separate transfer of H+ and NH3 on a unique two-lane pathway following NH4+ sequestration and deprotonation. This mechanism ensures that ammonium – which occurs mainly in protonated form in the aqueous phase – is efficiently translocated across the membrane, while maintaining strict selectivity against K+, a monovalent cation of similar size. This previously unobserved principle is likely to form a new paradigm for the electrogenic members of the Amt/Mep/Rh family. Similar mechanisms may be utilized by other membrane transporters to facilitate the selective translocation of pH-sensitive molecules.
Results and discussion
AmtB and NeRh50 activity is electrogenic
Motivated by our finding that the activity of Escherichia coli AmtB is electrogenic (Mirandela et al., 2019), we first investigated the transport mechanism of the Rh50 protein from Nitrosomonas europaea (NeRh50). Rh and Amt proteins are distant homologs, and thus a functional distinction between both subfamilies has been proposed (Huang and Ye, 2010). The architecture of NeRh50 is highly representative of Rh proteins (Gruswitz et al., 2010; Lupo et al., 2007) which have been repeatedly reported to serve as electroneutral NH3 or CO2 gas channels (Cherif-Zahar et al., 2007; Hub et al., 2010a; Li et al., 2007; Lupo et al., 2007; Weidinger et al., 2007). The activity of purified NeRh50 reconstituted into liposomes was quantified using Solid-Supported Membrane Electrophysiology (SSME) (Bazzone et al., 2017) experiments, where we recorded a NH4+-selective current (Figure 1) with a decay rate that is strongly dependent on the lipid-to-protein ratio (LPR; Table 1, Figure 1—figure supplement 1). Expressed in a Saccharomyces cerevisiae triple-mepΔ strain, deprived of its three endogenous Mep ammonium transporters, NeRh50 complemented the growth defect on minimal medium containing ammonium as sole nitrogen source (Figure 1). The electrogenic transport activity observed for NeRh50 and AmtB may suggest a common transport mechanism amongst the distant Amt and Rh proteins, but more experiments are needed to conclusively confirm this. Also, the NH4+ selectivity of both transporters further highlighted the question of how these proteins achieve selective charge translocation through their hydrophobic pore.
Figure 1 with 1 supplement see all
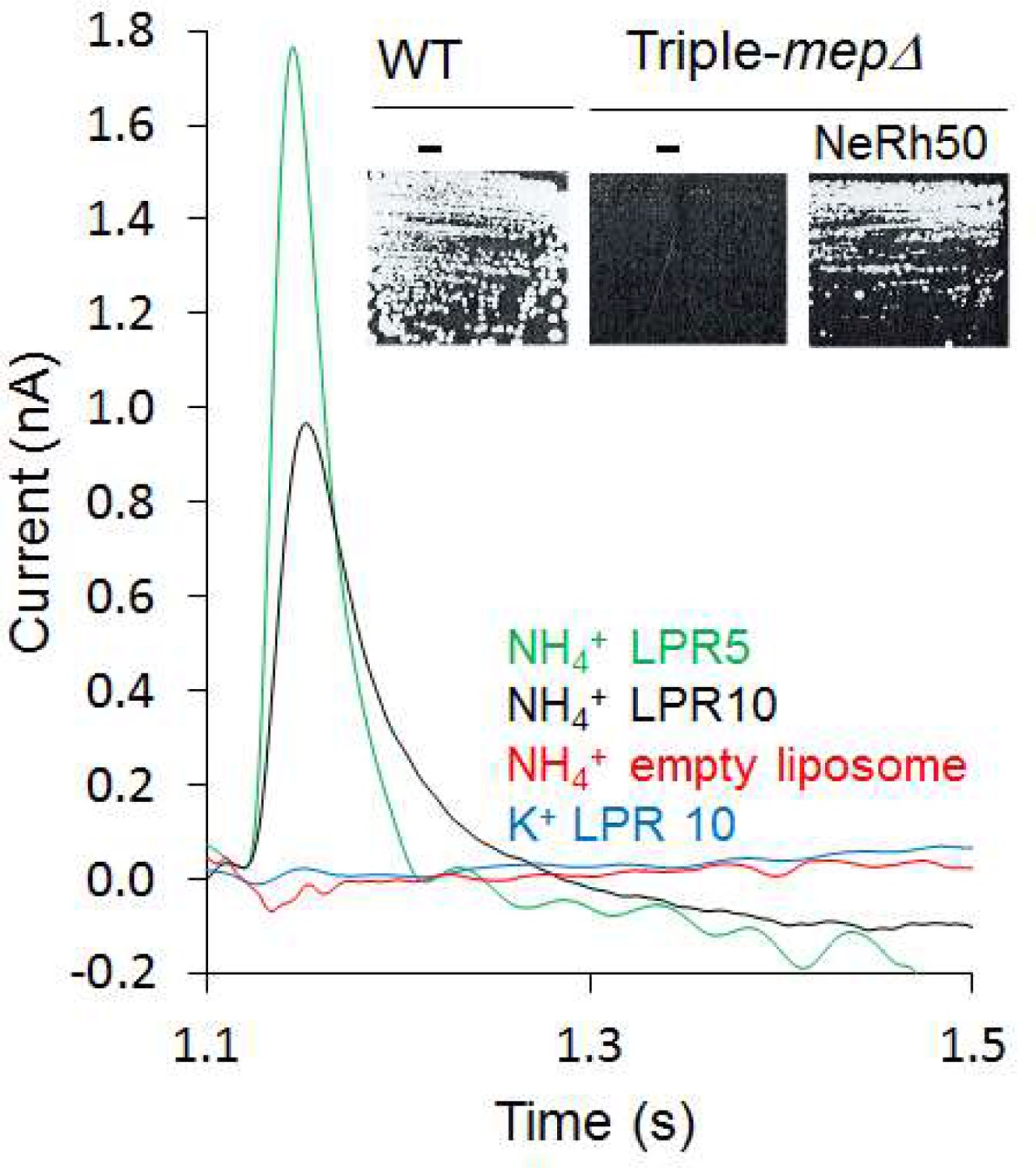
Characterization of the activity of NeRh50.
Transient current measured using SSME after a 200 mM pulse (ammonium or potassium). Insert: Yeast complementation by NeRh50 (strain 31019b, mep1Δ mep2Δ mep3Δ ura3) on minimal medium supplemented with 3 mM ammonium as sole nitrogen source.
-
Figure 1—source data 1
Characterization of the activity of NeRh50.
- https://cdn.elifesciences.org/articles/57183/elife-57183-fig1-data1-v3.xlsx
Table 1
Decay time constants (s−1) of transient currents triggered after an ammonium or potassium pulse of 200 mM in proteoliposomes containing AmtB at various LPR*.
| NH4+ | K+ | |||
|---|---|---|---|---|
| Variant | LPR 10 | LPR 5 | LPR 10 | LPR 5 |
| AmtB-WT | 13.4 ± 1.5 | 18.7 ± 1.0 | NC | NC |
| D160A | 21.6 ± 1.2 | 24.3 ± 1.5 | NC | NC |
| D160E | 17.03 ± 2.84 | 19.53 ± 1.8 | NC | NC |
| H168A H318A | 29.5 ± 2.1 | 29.8 ± 2.6 | NC | NC |
| S219A H168A H318A | NC | NC | NC | NC |
| H168A | 28.3 ± 1.5 | 38.0 ± 1.0 | 2.7 ± 0.5 | 5.2 ± 1.0 |
| H318A | 22.56 ± 2.63 | 28.25 ± 3.1 | 10.07 ± 1.7 | 15.64 ± 2.1 |
| NeRh50 | 24.0 ± 1.7 | 39.0 ± 3.6 | NC | NC |
-
*NC: No transient current recorded.
-
Table 1—source data 1
Decay time constants (s−1) of transient currents triggered after an ammonium or potassium pulse of 200 mM measured by SSME.
- https://cdn.elifesciences.org/articles/57183/elife-57183-table1-data1-v3.xlsx
Two interconnected water wires form an H+ translocation pathway in AmtB
We next made use of the most substantive body of structural information available for the archetypal ammonium transporter AmtB from E. coli and its variants to decipher the molecular mechanism of electrogenic NH4+ transport (Dias Mirandela et al., 2018). Computational (Wang et al., 2012) and experimental studies (Ariz et al., 2018) have suggested that deprotonation of NH4+ is likely to be a major step in ammonium transport. We therefore aimed to identify dynamic polar networks across AmtB that could form a transfer pathway through the protein for the translocation of H+, coming from NH4+ deprotonation. AmtB forms homotrimers in the cytoplasmic membrane, in which each monomer exhibits a potential periplasmic NH4+ binding region (S1) near residue D160, followed by a strongly hydrophobic pore leading towards the cytoplasm (Figure 2A; Khademi et al., 2004). Two highly conserved histidine residues, H168 and H318, protrude into the lumen, forming the family’s characteristic ‘twin-His’ motif (Javelle et al., 2006). The only variation in the twin-His motif in members of the Amt/Mep/Rh family is in numerous fungal Mep transporters where the first His, corresponding to H168, is replaced by a Glu (Javelle et al., 2006). The general conservation pattern in the AmtB pore, as analysed with ConSurf (Ashkenazy et al., 2016), is shown in Figure 2—figure supplement 1.
Figure 2 with 4 supplements see all
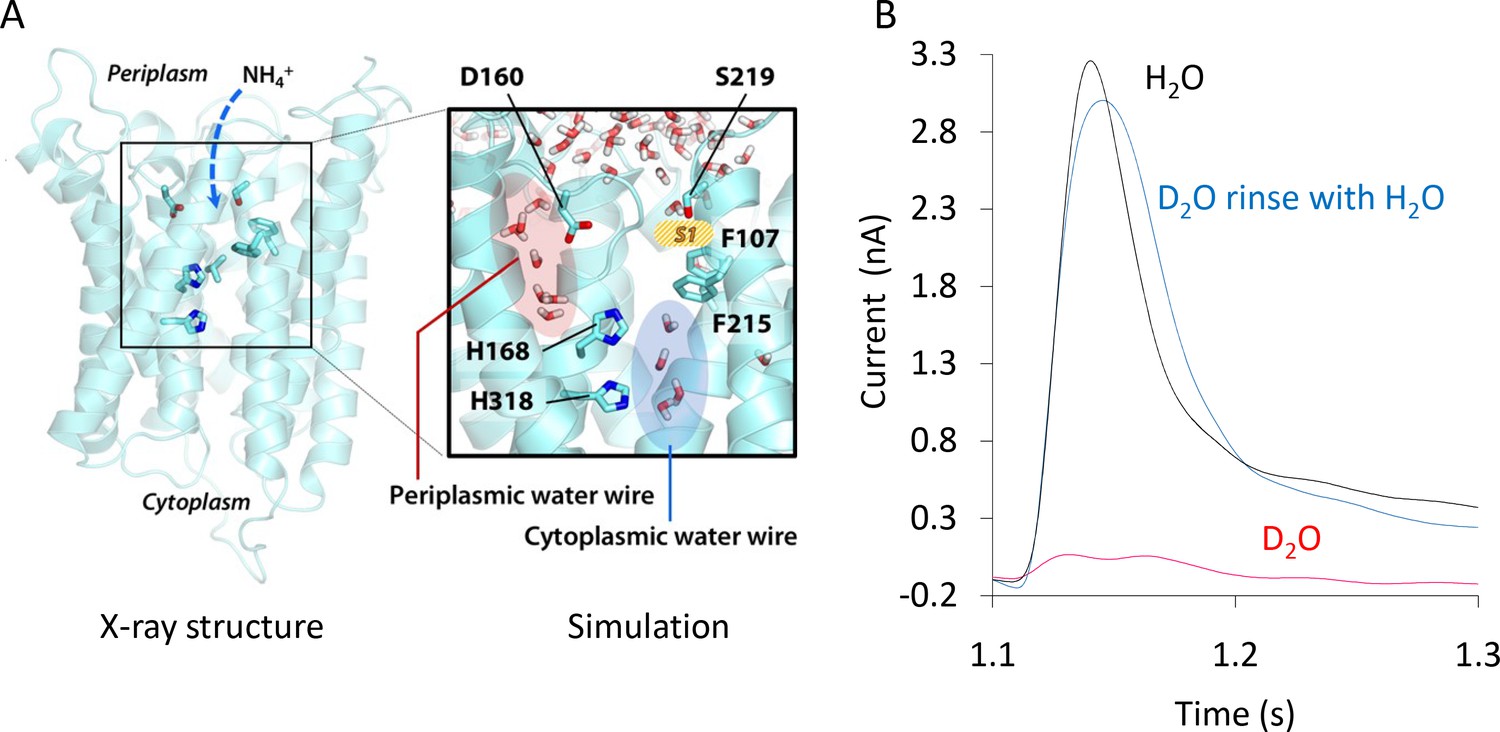
Formation and functionality of the periplasmic (PWW) and cytoplasmic (CWW) water wires in AmtB.
(A) Extended atomistic simulations show a hydration pattern across the protein, in which cytoplasmic and periplasmic water wires, connected via H168, form a continuous pathway for proton transfer from the S1 NH4+ sequestration region to the cytoplasm. (B) Transient currents measured following a 200 mM ammonium pulse on sensors prepared with solutions containing either H2O (black) or D2O (red). D2O sensors were rinsed with H2O solutions and subsequently exposed to another 200 mM ammonium pulse (blue).
-
Figure 2—source data 1
Functionality of the periplasmic (PWW) and cytoplasmic (CWW) water wires in AmtB.
- https://cdn.elifesciences.org/articles/57183/elife-57183-fig2-data1-v3.xlsx
To locate potential polar transfer routes, we performed atomistic molecular dynamics (MD) simulations of AmtB in mixed lipid bilayers. The simulations initially showed hydration of part of the putative hydrophobic NH3 pathway from the twin-His motif to the cytoplasm (cytoplasmic water wire – CWW; Figure 2A), confirming previous observations (Lamoureux et al., 2007). Notably, a new observation we made over longer simulation timescales is the presence of a previously unidentified second water-filled channel (periplasmic water wire - PWW). The PWW spans from residue D160 near the S1 region to the central twin-His motif (Figure 2A) and is formed both in simulations without and with applied membrane voltage Vm (Figure 2A - Figure 2—figure supplement 2; Vm in E. coli ~ −140 mV [Felle et al., 1980]).
As the protonation pattern of the twin-His motif has been found to play a role in the hydration of the protein (Ishikita and Knapp, 2007), two different tautomeric states of the twin-His motif were systematically probed in the simulations. The tautomeric state in which H168 is protonated on its Nδ and H318 is protonated on its Nε atom is referred to as ’DE’, while ’ED’ terms the twin-His configuration where H168 is protonated on Nε and H318 is protonated on Nδ (Figure 2A- Figure 2—figure supplement 2, 3, 4). Formation of the CWW is observed to occur within a few nanoseconds at the beginning of each simulation. In the DE tautomeric state, the cytoplasmic pocket of each subunit almost continuously remains occupied by 3–4 water molecules for the rest of the simulation (Figure 2A - Figure 2—figure supplement 2, 3, 4; data for 0 mV membrane voltage). In the ED state, greater fluctuations in the number of water molecules in the chain are seen, and the average occupancy is decreased. Using a cut-off value of three water molecules per subunit, a complete water chain is present during 79% of the simulations in the DE state, and only during 12% of the simulated time in the ED state. The PWW is generally more transiently occupied than the cytoplasmic channel; however, we record up to 23% occupancy with at least three water molecules when the histidine sidechains are in the ED tautomeric state (Figure 2A - Figure 2—figure supplement 2, 3, 4).
Both water wires are connected via the twin-His motif, which bridges the aqueous chains, while preventing the formation of a continuous water channel in the simulations. Although neither the CWW nor the PWW are sufficiently wide to allow the transfer of solvated NH4+, water molecules and histidine side chains could serve as efficient pathways to facilitate proton transfer in proteins (Acharya et al., 2010). As shown in Figure 2—figure supplement 1, the key residues that line both water wires in AmtB are highly conserved in the family.
The interconnected water wires are functionally essential to AmtB activity
To experimentally test if the water wires are essential for proton conduction during the AmtB transport cycle, we made use of the reduced deuteron mobility of heavy water D2O. Because deuterons have twice the mass of a proton and the bond strength is increased, the deuteron mobility is reduced by 30% for each D2O molecule compared to normal water (Wiechert and Beitz, 2017). Since the polar network of water we identified involves more than three water molecules (Figure 2A), AmtB should be nearly inactive if tested in the presence of D2O. Indeed, we found that in an SSME-based assay where all buffers used to prepare the proteoliposomes and SSM sensors were made using D2O, AmtB activity was completely abolished compared to buffer containing water (Figure 2B). After rinsing the sensor prepared in D2O with water, AmtB re-gained 100% of its activity measured by SSME, showing that the presence of D2O did not affect the protein itself or the integrity of the proteoliposomes (Figure 2B). Further calculations suggested that H+ transfer between the water molecules is possible both within the PWW and CWW and could occur with high rates (the highest energy barrier is ~18 kJ/mol in the cytoplasmic wire near the twin-His motif; Table 2). Taken together, the experimental and computational data suggest that proton transfer between water molecules, most likely the PWW and CWW detected in the simulations, may underpin the electrogenic activity of AmtB.
Table 2
Free energies for proton translocation through the cytoplasmic and periplasmic water wires and neighboring water molecules (bulk)*.
| Z (Å) | Free energy (kJ/mol) | |||
|---|---|---|---|---|
| (bulk) | Peripl. water wire | wat1 | 14.7 | 0.0 |
| wat2 | 12.7 | 8.7 | ||
| wat3 | 10.7 | 15.0 | ||
| wat4 | 8.3 | 14.4 | ||
| wat5 | 6.1 | 7.5 | ||
| D160 | wat6 | 5.4 | 11.0 | |
| wat7 | 3.2 | 14.4 | ||
| wat8 | 0.6 | 18.5 | ||
| H168 | ||||
| cytopl. water wire | wat9 | −0.4 | 17.3 | |
| wat10 | −0.8 | 14.4 | ||
| wat11 | −3.2 | 12.1 | ||
| H318 | wat12 | −5.1 | 13.8 | |
-
*The vertical coordinate z was calculated relative to the position of the sidechain of H168. Positions of the sidechains of D160, H168 and H318 with respect to the periplasmic and cytoplasmic water wires are indicated in the left column.
AmtB activity is not driven by the proton motive force
In the absence of ammonium, a proton pulse did not trigger a discernible current and additionally, in the presence of ammonium, an inward-orientated pH gradient did not increase AmtB activity (Figure 3). These findings suggest that there is no H+-dependent symport activity of AmtB, showing instead that AmtB is not able to translocate a proton in the absence of NH4+, and indicating that the current induced by AmtB activity is generated by specific deprotonation of the substrate and subsequent H+ translocation. Furthermore, they show that AmtB cannot act as an uncoupler, which raises the question of proton selectivity and the coupling between NH3 and H+ transfer (Boogerd et al., 2011; Maeda et al., 2019). Our current data suggest that the PWW is transiently occupied and that its occupancy is strongly dependent on the particular state and conformation of D160, since even a D to E conservative change abolished presence of the PWW (Figure 4A). Any disruption of the PWW will, in turn, impede the capability of AmtB to transfer H+. The functionally relevant conformation and protonation state of D160 that stabilize the PWW is likely to be coupled to the presence of a charged substrate binding near S1, thereby linking substrate binding and deprotonation to H+ transfer.
Figure 3
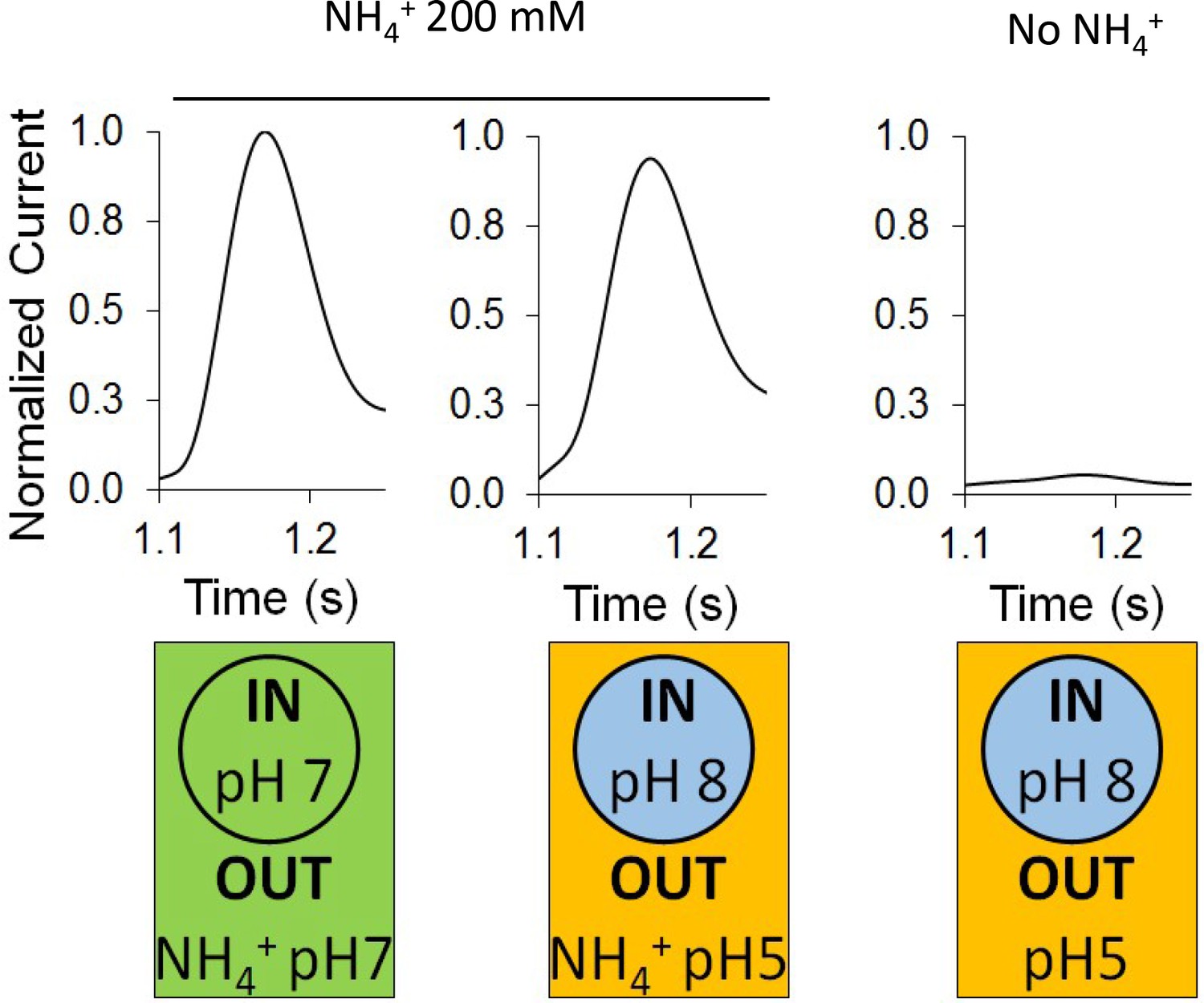
Effect of a proton gradient on AmtB activity.
The transient currents were measured using SSME following an ammonium pulse of 200 mM at pH 7 (left) or under an inwardly directed pH gradient in the presence (center) or absence (right) of ammonium. eight sensors from two independent protein purification batches were measured, with three measurements recorded for each sensor. Single representative traces were chosen to visualize the results. Each sensor was measured in the order pH (in/out) 7/7, 8/5, 8/5 (this time without NH4+), and finally 7/7 again to be sure that the signals do not significantly decrease with time. The data are normalized against the measurements done at pH7 in/out for each sensor.
-
Figure 3—source data 1
Effect of a proton gradient on AmtB activity.
- https://cdn.elifesciences.org/articles/57183/elife-57183-fig3-data1-v3.xlsx
Figure 4 with 2 supplements see all
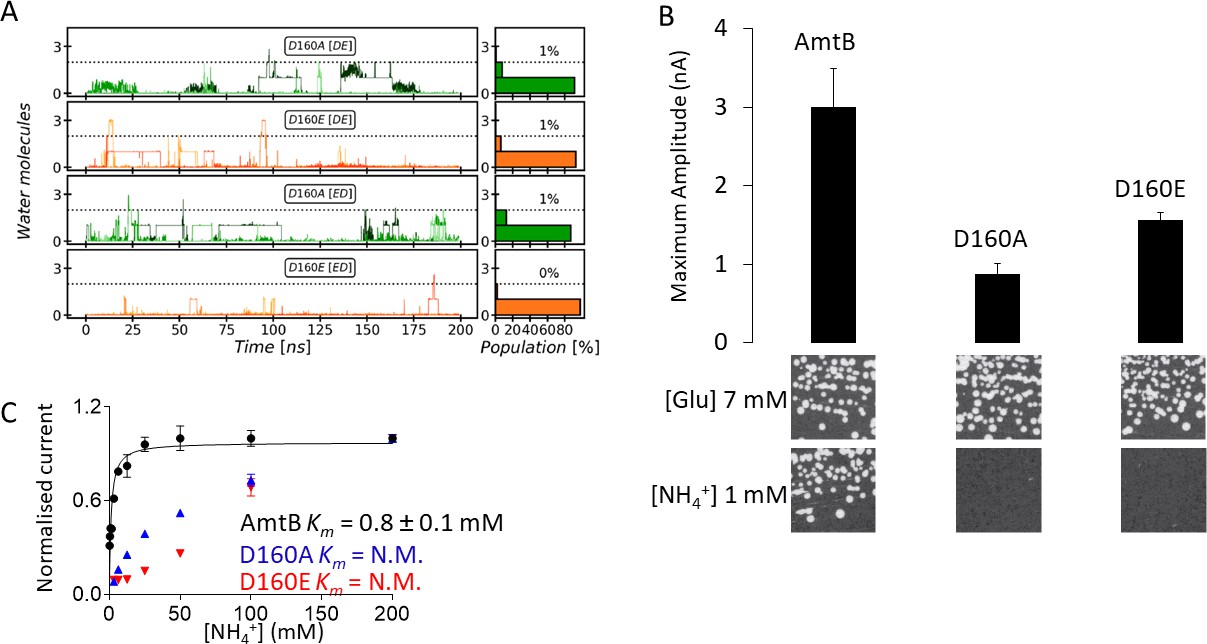
Effect of D160 substitutions.
(A) The Periplasmic Water Wire (PWW) in the D160A and D160E variants. We observe no significant occupancy of the PWW above the threshold of at least three water molecules in the D160A and D160E AmtB variants, irrespective of the tautomeric protonation states of H168 and H318 (DE or ED, see Materials and method section). (B) Upper panel: maximum amplitude of the transient current measured using SSME following a 200 mM ammonium pulse. Eight sensors from two independent protein purification batches were measured, with three measurements recorded for each sensor (means ± SD). Lower panel: yeast complementation test (strain 31019b, mep1Δ mep2Δ mep3Δ ura3) using 7 mM Glutamate (Glu) or 1 mM ammonium as a sole nitrogen source. The growth tests have been repeated twice. (C) Kinetics analysis of the transport of ammonium. The maximum amplitudes recorded after a 200 mM ammonium pulse have been normalized to 1.0 for comparison. N.M.: Non Measurable. eight sensors from two independent protein purification batches were measured, with three measurements recorded for each sensor (means ± SD).
-
Figure 4—source data 1
Effect of D160 substitutions on AmtB activity measured by SSME.
- https://cdn.elifesciences.org/articles/57183/elife-57183-fig4-data1-v3.xlsx
The residue D160 is essential to stabilize the PWW
As the PWW is formed near the sidechain of D160, an invariant residue in the Amt/Mep/Rh superfamily (Marini et al., 2006; Thomas et al., 2000), we further investigated the role of this residue in ensuring PWW and CWW stability by simulating the AmtB D160A and D160E mutants. Both mutants were stable on the time scale of our simulations and we did not detect major rearrangements in the protein. Moreover, all the elution profiles of the purified WT and variants proteins obtained by analytical size exclusion chromatography, before and after solubilization of the proteoliposomes in 2% DDM, were identical, showing a single monodisperse peak eluting between 10.4–10.6 ml (Figure 4—figure supplement 1). Taken together, these results suggest that major structural re-arrangements in the mutants are unlikely to occur. The simulations revealed no difference in the formation of the CWW in the D160A and D160E variants compared to the WT, however the formation of the PWW is almost completely abolished in the presence of these mutations (Figure 4A).
We then expressed wild-type AmtB as well as the D160A and D160E mutants in S. cerevisiae triple-mepΔ. Using ammonium as the sole nitrogen source, we found that cells expressing the mutants failed to grow, showing that AmtBD160A or AmtBD160E are unable to replace the function of the endogenous Mep transporters (Figure 4B).
The activity of the purified variants reconstituted into liposomes was next quantified using SSME. Electrogenic transport activity, triggered by a 200 mM ammonium pulse, led to a transient current with a maximum amplitude of 3.38 nA in AmtB, while AmtBD160A and AmtBD160E displayed reduced maximum currents of 0.63 nA and 1.42 nA respectively (Figure 4B, Figure 4—figure supplement 2). Importantly, the lifetime of currents in both variants was unaffected by changes in liposomal LPR, and therefore the small transient current accounts for the binding of a NH4+ to the proteins and not a full translocation cycle (Table 1, Figure 4—figure supplement 2; Bazzone et al., 2017). Additionally, it was impossible to determine with confidence a catalytic constant (Km) for both variants since no clear saturation was reached, even after an ammonium pulse of 200 mM (Figure 4C). These results thus demonstrate that AmtBD160A and AmtBD160E are transport-deficient. Our data show that residue D160 plays a central role in the transport mechanism as opposed to having a strictly structural role as previously suggested (Khademi et al., 2004). Moreover, the fact that the conservative D to E variation at position 160 impairs ammonium transport via AmtB indicates that D160 does not only show electrostatic interaction with NH4+ at the S1 site but is also involved in the translocation mechanism by stabilizing the PWW.
AmtB switches from transporter to channel-like activity in the absence of the ‘twin-His’ motif
The CWW and PWW are connected via the twin-His motif, which bridges the aqueous chains, while preventing the formation of a continuous water channel in the simulations (Figure 2A). We therefore next probed if the twin-His motif enables proton transfer between the two water wires by recording the activities of twin-His variants. Expressed in S. cerevisiae triple-mepΔ, AmtBH168A/H318A did not support cell growth on low ammonium (Figure 5A). In-vitro SSME measurements with this variant displayed LPR-independent current decay rates (Figure 5A, Figure 5—figure supplement 1, Table 1), showing that the residual current is caused by the association of NH4+ to AmtB without further transport. No current was recorded for the triple mutant AmtBS219A/H168A/H318A, in which binding at the periplasmic face was further altered, confirming that the residual current reflects NH4+ interaction near S1 (Figure 5—figure supplement 1). The double-His mutant AmtBH168A/H318A is thus able to interact with NH4+ but cannot transport the substrate across the membrane. This supports our previous structural analysis showing that the CWW in the pore of the double-His mutant AmtBH168A/H318A is absent (Javelle et al., 2006).
Figure 5 with 2 supplements see all
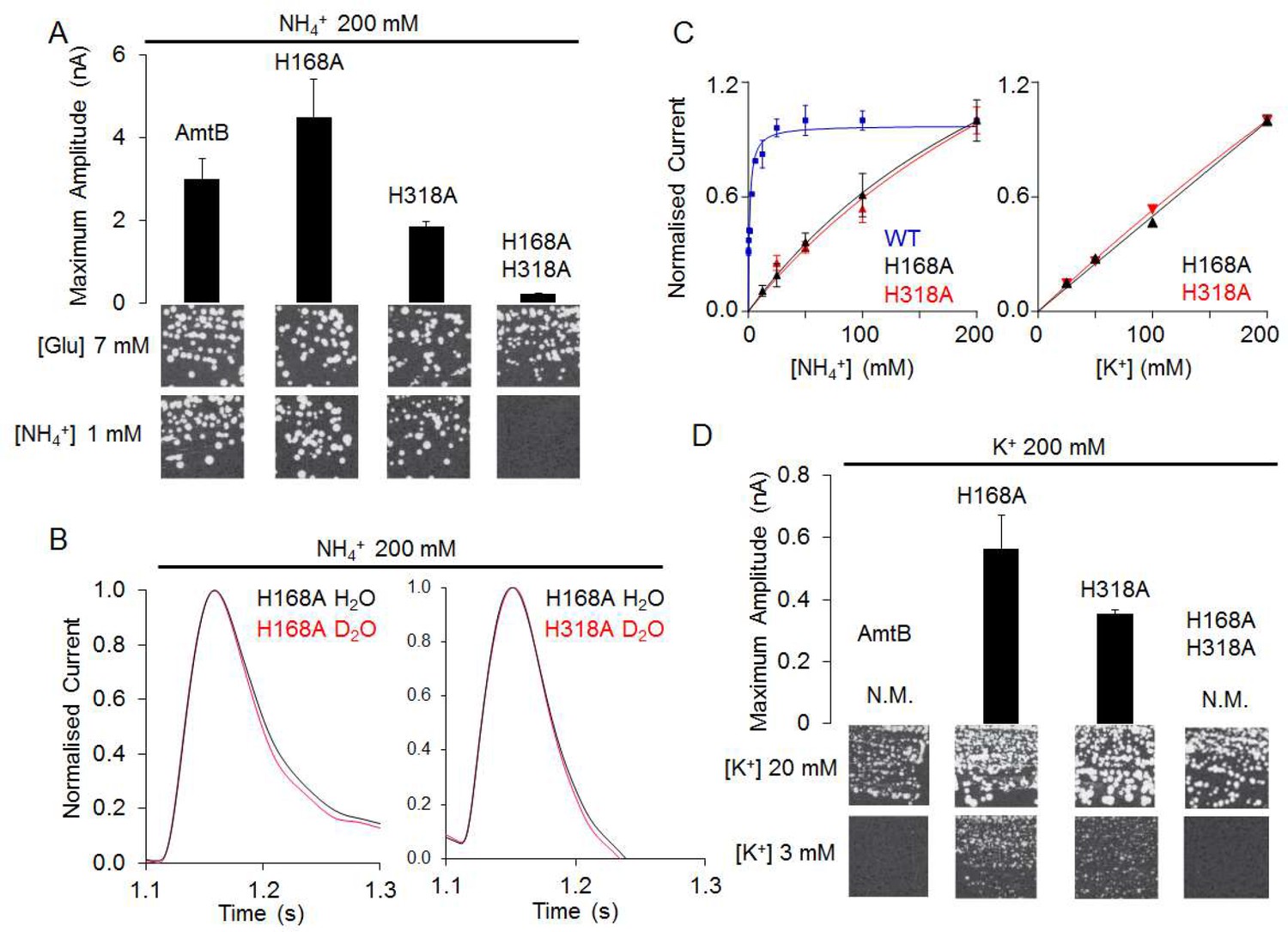
The AmtBH168A and AmtBH318A lose their specificity toward ammonium.
(A) Upper panels: maximum amplitude of the transient current measure using SSME after a 200 mM ammonium pulse. Eight sensors from two independent protein purification batches were measured, with three measurements recorded for each sensor (means ± SD). Lower panels: yeast complementation test (strain 31019b, mep1Δ mep2Δ mep3Δ ura3) using 7 mM Glutamate (Glu) or 1 mM ammonium as a sole nitrogen source. The growth tests have been repeated twice. (B) Transient currents measured using SSME following a 200 mM ammonium pulse on sensors prepared with solutions containing either H2O (black) or D2O (red). The maximum amplitudes recorded after a 200 mM ammonium pulse on sensor prepared in H2O have been normalized to 1.0 for comparison. eight sensors from two independent protein purification batches were measured, with three measurements recorded for each sensor (means ± SD). (C) Kinetics analysis of the transport of NH4+ (or K+ in AmtBH168A (black), AmtBH318A (red) and WT-AmtB (bleu, only for NH4+, as no signal was measurable with K+). The maximum amplitudes recorded after a 200 mM NH4+ or K+ pulse have been normalized to 1.0 for comparison. Eight sensors from two independent protein purification batches were measured, with three measurements recorded for each sensor (means ± SD). (D) Upper panels: maximum amplitude of the transient current measured using SSME after a 200 mM potassium pulse. N.M. Non Measurable. Eight sensors from two independent protein purification batches were measured, with three measurements recorded for each sensor (means ± SD). Lower panels: yeast complementation test (strain #228, mep1Δ mep2Δ mep3Δ trk1Δ trk2Δ leu2 ura3) using media supplemented with 20 mM or 3 mM KCl. The growth test has been repeated twice.
-
Figure 5—source data 1
Effect of H168 and/or H318 substitution on AmtB activity and selectivity measured by SSME.
- https://cdn.elifesciences.org/articles/57183/elife-57183-fig5-data1-v3.xlsx
By contrast, the two single histidine-to-alanine substitutions in the twin-His motif unexpectedly produced an LPR-dependent current in our SSME recordings (Figure 5A, Figure 5—figure supplement 1, Table 1). Furthermore, triple-mepΔ yeast cells expressing these variants were able to grow in the presence of low ammonium concentrations (Figure 5A). Our previous crystal structure (Javelle et al., 2006) and our MD simulations (Figure 5—figure supplement 2) show increased hydration in the area around A168, which could potentially form a pathway for direct translocation of NH4+ without a deprotonation step. To test this hypothesis, we measured the activity of AmtBH168A and AmtBH318A in D2O conditions, as described above. Crucially, the activity of both variants measured in the presence or absence of D2O was similar (Figure 5B), in contrast to native AmtB where no activity was recorded in D2O (Figure 2B), showing that proton transfer between water molecules is not a key mechanistic feature in the activity of the mutants. Additionally, the translocation of NH4+ is not saturable in the tested concentration range [12.5–200 mM] for AmtBH168A and AmtBH318A (Figure 5C). Summarizing, these results suggest that AmtB switches from transporter- to channel-like activity in the absence of the twin-His motif, directly translocating hydrated NH4+ through the pore. In the wild-types of Amt/Mep/Rh protein family members, the twin-His motif is highly conserved, which shows that transporter, as opposed to channel activity, is mechanistically crucial for the function of these proteins. The only variation seen in naturally occurring sequences is a replacement of the first His by Glu in some fungal Mep proteins (Javelle et al., 2006; Thomas et al., 2000). Channel activity is so far only observed for the alanine mutants, not the wild-type. We hypothesized that transport activity might thus be key to ensure ion selectivity of AmtB, since NH4+ and K+ are cations of similar size and hydration energy (Aydin et al., 2020).
The twin-His motif interconnects the two water wires to ensure the selectivity of AmtB
Since NH4+ was directly translocated in the absence of the twin-his motif and earlier studies implicated a role of the motif in AmtB selectivity (Ganz et al., 2020; Hall and Yan, 2013), we repeated our SSME experiments on the AmtBH168A and AmtBH318A variants using the competing K+ ion as substrate. A 200 mM K+ pulse triggered currents in both variants, whose decay rates strongly depended on the LPR (Figure 5D, Table 1, Figure 5—figure supplement 1). Furthermore, the single His variants, but not native AmtB, complemented the growth defect of a yeast strain lacking its three endogenous ammonium (Mep) and potassium (Trk) transporters when a limited concentration of K+was present (Figure 5D). The K+ translocation activity is not saturable in the substrate range [12.5–200 mM] (Figure 5C). These results demonstrate that both variants, AmtBH168A and AmtBH318A, translocate K+ ions across the membrane. The substitutions within the twin-His motif thus abolished selectivity for NH4+.
The presence of both histidine residues is therefore critical in ammonium transport, since permeability of ammonium transporters for K+ would compromise ionic homeostasis and disrupt the membrane potential of E. coli cells, which crucially depends on maintaining K+ concentration gradients across the membrane. Moreover, since AmtB is expressed in E. coli under nitrogen starvation conditions (low NH4+/K+ ratio), loss of selectivity for NH4+ would impede ammonium uptake. Our results thus demonstrate that the twin-His motif, which is highly conserved amongst members of the family, is an essential functional element in the transport mechanism, preventing the transport of competing cations, whilst providing a pathway for proton transfer by bridging the periplasmic and the cytoplasmic water wires.
NH3 permeation through the hydrophobic pore
Umbrella sampling free-energy calculations were performed to establish the rate limiting step of NH4+ transport. Our calculations show that NH3 translocation experiences only a moderate energy barrier (~10 kJ/mol) at the periplasmic hydrophobic constriction region (F107 and F215) (Figure 6). The starting points of the sampling windows were determined from the centers of HOLE calculations (Smart et al., 1996), optimizing the pathway of NH3 translocation across the pore. A possible influence from this selection regarding the pathway was further reduced by allowing the molecule to move freely perpendicular to the pore axis within a radius of 5 Å in addition to extensive sampling; however, residual bias from window selection cannot be completely excluded. From the free energy profile of NH3 translocation, we identified a shallow binding site below the twin-His motif (~5 kJ/mol). This is followed by a second hydrophobic region (I28, V314, F31 and Y32) that forms a small energy barrier between this binding site and the cytoplasmic exit. The increased residence time of NH3 within this energy minimum suggests that reprotonation to NH4+, caused by the cytoplasmic pH, occurs in this region (Figure 6). Since both energy barriers for H+ transfer along the water chains and NH3 permeation are relatively small, we concluded that either initial deprotonation or proton transfer across the twin-His motif could be rate-limiting for overall NH4+ transport.
Figure 6
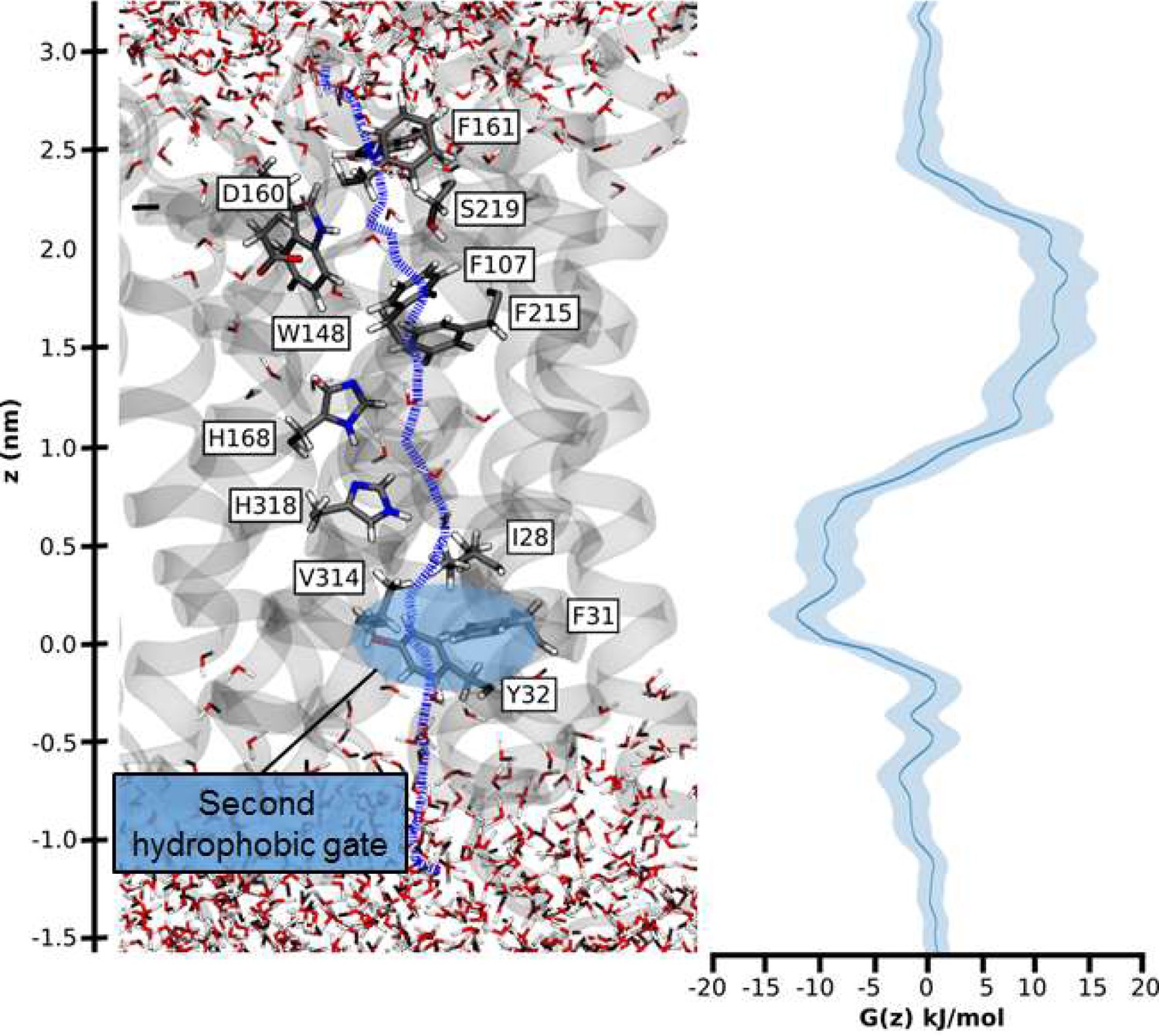
Hydrophobic pathway and energetics for NH3 translocation in AmtB.
We probed an optimal pathway for NH3 transfer during our PMF calculations (left, purple dash trajectory) in the presence of both the PWW and CWW. The software HOLE (49) was used to determine the most likely transfer route. The pathway from the periplasm to the cytoplasm traverses the hydrophobic gate region (F107 and F215), crosses the cavity next to the twin-His motif (H168 and H318) occupied by the CWW, and continues across a second hydrophobic region (I28, V314, F31, Y32) before entering the cytoplasm.
Conclusion
A new model for the mechanism of electrogenic ammonium transport therefore emerges from our findings (Figure 7). After deprotonation of NH4+ at the periplasmic side, a previously undiscovered polar conduction route enables H+ transfer into the cytoplasm. A parallel pathway, lined by hydrophobic groups within the protein core, facilitates the simultaneous transfer of uncharged NH3, driven by concentration differences. On the cytoplasmic face, the pH of the cell interior leads to recombination to NH4+, most likely near a second hydrophobic gate (Figure 6). The twin-His motif, which bridges the water chains constitutes the major selectivity gate for NH4+ transport preventing K+ flow. We propose that this mechanism is conserved amongst the electrogenic members of the Amt/Mep/Rh family. Importantly, two RhAG polymorphisms associated to the overhydrated stomatocytosis human syndrome have also acquired the ability to transport K+. Thus, deciphering the transport mechanism of two archetypal members of the family such as AmtB and NeRh50 could bring new insights to the understanding of substrate specificity determinants in Rh proteins in the context of human diseases (Bruce et al., 2009).
Figure 7
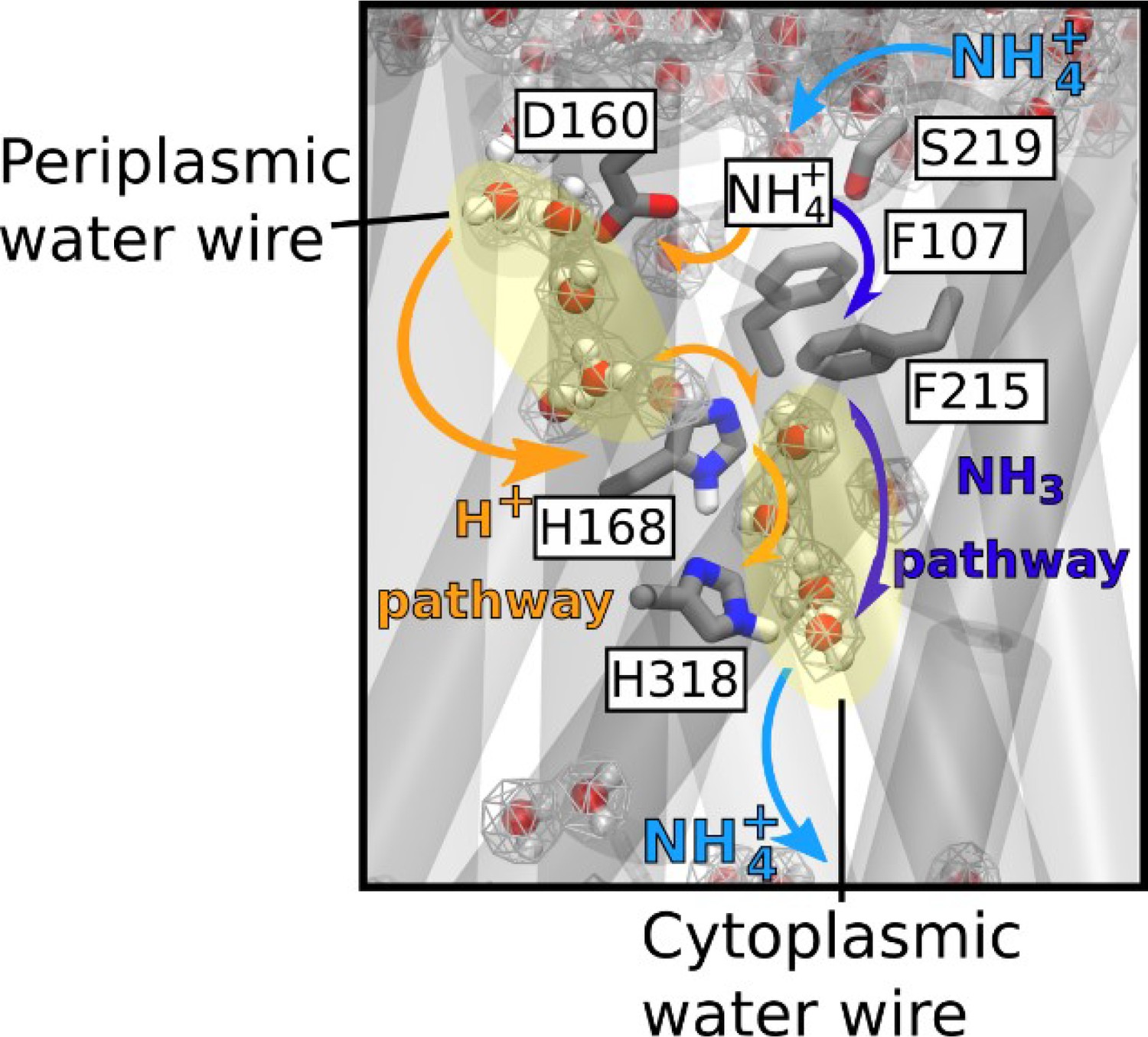
Mechanism of electrogenic NH4+ translocation in AmtB.
Following sequestration of NH4+ at the periplasmic face, NH4+ is deprotonated and H+ and NH3 follow two separated pathways to the cytoplasm (orange arrows depict the pathway for H+ transfer, dark blue arrows for NH3), facilitated by the presence of two internal water wires. NH3 reprotonation likely occurs near the cytoplasmic exit (Figure 6). The hydrated regions within the protein as observed in simulations are highlighted by wireframe representation, crucial residues involved in the transport mechanism are shown as sticks.
Our findings define a new mechanism, by which ionizable molecules that are usually charged in solution are selectively and efficiently transported across a highly hydrophobic environment like the AmtB/Rh pore. Alongside size-exclusion and ion desolvation (Kopec et al., 2018), it adds a new principle by which selectivity against competing ions can be achieved. Other biological transport systems, like the formate/nitrite transporters, may share similar mechanisms involving deprotonation-reprotonation cycles (Wiechert and Beitz, 2017).
Materials and methods
Key resources table
| Reagent type (species) or resource | Designation | Source or reference | Identifiers | Additional information |
|---|---|---|---|---|
| Gene (Escherichia coli) | AmtB | Zheng et al., 2004 | Uniprot: C3TLL2 | |
| Gene (Nitrosomonas europea) | Rh50 | Lupo et al., 2007 | Uniprot Q82 × 47 | |
| Strain, strain background (Escherichia coli) | C43 (DE3) | Miroux and Walker, 1996 | Chemically competent cells | |
| Strain, strain background (Escherichia coli) | GT1000 | Javelle et al., 2004 | Chemically competent cells | |
| Recombinant DNA reagent | pET22b (+) | Novagen | Cat# - 69744 | |
| Recombinant DNA reagent | pDR195 | Rentsch et al., 1995 | Addgene - 36028 | High copy yeast expression vector |
| Recombinant DNA reagent | pAD7 | Cherif-Zahar et al., 2007 | pESV2-RH50(His)6 | |
| Recombinant DNA reagent | p426MET25 | Mumberg et al., 1994 | ||
| Recombinant DNA reagent | PZheng | Zheng et al., 2004 | pET22b-AmtB(His)6 | |
| Recombinant DNA reagent | pGDM2 | This study | pET22b-AmtB(His)6H168AH318A | |
| Recombinant DNA reagent | pGDM4 | This study | pET22b-AmtB(His)6D160A | |
| Recombinant DNA reagent | pGDM5 | This study | pET22b-AmtB(His)6D160E | |
| Recombinant DNA reagent | pGDM6 | This study | pET22b-AmtB(His)6S219AH168AH318A | |
| Recombinant DNA reagent | pGW2 | This study | pET22b-AmtB(His)6H168A | |
| Recombinant DNA reagent | pGDM9 | This study | pDR195-AmtB(His)6D160A | |
| Recombinant DNA reagent | pGDM10 | This study | pDR195-AmtB(His)6D160E | |
| Recombinant DNA reagent | pGDM12 | This study | pDR195-AmtB(His)6H168AH318A | |
| Recombinant DNA reagent | pGDM13 | This study | pDR195-AmtB(His)6S219AH168AH318A | |
| Recombinant DNA reagent | pGW7 | This study | pDR195-AmtB(His)6H168A | |
| Sequence-based reagent | AmtBS219A F | IDT | PCR Primer (Mutagenesis) | GGTGGCACCGTGGTGGATATTAACGCCGCAATC |
| Sequence-based reagent | AmtBD160A F | IDT | PCR Primer (Mutagenesis) | CTCACGGTGCGCTGGCCTTCGCGGGTGGCACC |
| Sequence-based reagent | AmtBD160E F | IDT | PCR Primer (Mutagenesis) | CTCACGGTGCGCTGGAGTTCGCGGGTGGCACC |
| Sequence-based reagent | AmtBH168A F | IDT | PCR Primer (Mutagenesis) | GGTGGCACCGTGGTGGCCATTAACGCCGCAATC |
| Sequence-based reagent | AmtBH318A F | IDT | PCR Primer (Mutagenesis) | TGTCTTCGGTGTGGCCGGCGTTTGTGGCATT |
| Sequence-based reagent | AmtB XhoI | IDT | PCR primer | AGTCCTCGAGATGAAGATAGCGACGATAAAA |
| Sequence-based reagent | AmtB BamHI | IDT | PCR primer | AGTCGGATCCTCACGCGTTATAGGCATTCTC |
| Sequence-based reagent | P5’NeRh | IDT | PCR primer | GCCACTAGTATGAGTAAACACCTATGTTTC |
| Sequence-based reagent | P3’NeRh | IDT | PCR primer | GCCGAATTCCTATCCTTCTGACTTGGCAC |
| Peptide, recombinant protein | AmtB(His)6 | This study | purified from E. coli C43 (DE3) cells | |
| Peptide, recombinant protein | AmtB(His)6D160A | This study | purified from E. coli C43 (DE3) cells | |
| Peptide, recombinant protein | AmtB(His)6 D160E | This study | purified from E. coli C43 (DE3) cells | |
| Peptide, recombinant protein | AmtB(His)6 H168AH318A | This study | purified from E. coli C43 (DE3) cells | |
| Peptide, recombinant protein | AmtB(His)6 S219AH168AH318A | This study | purified from E. coli C43 (DE3) cells | |
| Peptide, recombinant protein | AmtB(His)6 H168A | This study | purified from E. coli C43 (DE3) cells | |
| Peptide, recombinant protein | AmtB(His)6 H318A | This study | purified from E. coli C43 (DE3) cells | |
| Peptide, recombinant protein | NeRh50(His)6 | This study | purified from E. coli C43 GT1000 cells | |
| Peptide, recombinant protein | XhoI | Promega | Cat# - R6161 | |
| Peptide, recombinant protein | BamHI | Promega | Cat# - R6021 | |
| Commercial assay or kit | Quikchange XL site-directed mutagensis kit | Agilent Technologies | Cat# 200516 | |
| Chemical compound, drug | n-dodecyl-β-D-maltopyranoside (DDM) | Avanti | Cat#- 850520 | |
| Chemical compound, drug | lauryldecylamine oxide (LDAO) | Avanti | Cat#- 850545 | |
| Chemical compound, drug | E. coli Polar Lipids | Avanti | Cat#−100600 | |
| Chemical compound, drug | Phosphotidylcholine (POPC) | Avanti | Cat#−850457 | |
| Software, algorithm | Graphpad Prism software | GraphPad Prism (https://www.graphpad.com) | Version 6.01 | |
| Software, algorithm | Origin Pro Software | Origin Labs (https://www.originlab.com) | Origin 2017 Version 94E | |
| Software, algorithm | SURFE2R Control Software | Nanion (https://www.nanion.de/en/) | V1.5.3.2 |
Mutagenesis
AmtB mutants were generated using the Quikchange XL site-directed mutagenesis kit (Agilent Technologies), according to the manufacturer’s instructions. The primers used for mutagenesis are listed in Key resources table. The template was the amtB gene cloned into the plasmid pET22b(+), as previously described (Zheng et al., 2004; Key resources table).
AmtB and NeRh50 expression in yeast and complementation test
Request a detailed protocolThe different variants of amtB were amplified using amtB cloned into pET22b(+) (Key resources table) as a template with the primers AmtB XhoI and AmtB BamHI (Key resources table) and then sub-cloned into the plasmids pDR195 (Key resources table). The NeRh50 gene was amplified from N. europaea genomic DNA (kind gift from Daniel J. Arp and Norman G. Hommes, Department of Botany and Plant Pathology, Oregon State University, Corvallis, USA) using the primers P5’NeRh and P3’NeRh (Key resources table), and was then cloned into the SpeI and EcoRI restriction sites of the high-copy vector p426Met25 (Key resources table), allowing controlled-expression of NeRh50 by the yeast methionine repressible MET25 promoter.
Saccharomyces cerevisiae strains used in this study are the 31019b strain (mep1Δ mep2Δ mep3Δ ura3) and the #228 strain (mep1Δ mep2Δ mep3Δ trk1Δ trk2Δ leu2 ura3) (Hoopen et al., 2010; Marini et al., 1997). The plasmids used in this study are listed in Key resources table. Cell transformation was performed as described previously (Gietz et al., 1992). For growth tests on limiting ammonium concentrations, yeast cells were grown in minimal buffered (pH 6.1) medium and for growth tests on limiting potassium concentrations, a minimal buffered (pH 6.1) medium deprived of potassium salts was used (Jacobs et al., 1980). 3% glucose was used as the carbon source and, 0.1% glutamate, 0.1% glutamine or (NH4)2SO4 at the specified concentrations were used as the nitrogen sources.
All growth experiments were repeated at least twice.
Protein purification
Request a detailed protocolAmtB(His6) cloned into the pET22b(+) vector (Key resources table) was overexpressed and purified as described previously (Zheng et al., 2004). The plasmid pAD7 (Key resources table) was used to overexpress NeRh50 in the E. coli strain GT1000 (Javelle et al., 2004). GT1000 was transformed with pAD7 and grown in M9 medium (Elbing and Brent, 2002), in which ammonium was replaced by 200 μg/ml glutamine as sole nitrogen source. NeRh50 was purified as described by Lupo et al., 2007 with minor modifications, namely: the membrane was solubilized using 2% lauryldecylamine oxide (LDAO) instead of 5% n-octyl-β-D-glucopyranoside (OG), and 0.09% LDAO was used in place of 0.5% β-OG in the final size exclusion chromatography buffer (50 mL Tris pH 7.8, 100 mL NaCl, 0.09% LDAO).
AmtB and NeRh50 insertion into proteoliposomes
Request a detailed protocolAmtB and NeRh50 were inserted into liposomes containing E. coli polar lipids/phosphatidylcholine (POPC) 2/1(wt/wt) as previously described (Mirandela et al., 2019). For each AmtB variant, proteoliposomes were prepared at lipid-to-protein ratios (LPRs) of 5, 10, and 50 (wt/wt). The size distribution of proteoliposomes was measured by dynamic light scattering (DLS) using a Zetasizer Nano ZS (Malvern Instruments, Malvern, UK). This analysis showed that the proteoliposomes had an average diameter of 110 nm (Figure 8). Proteoliposomes were divided into 100 µL aliquots and stored at −80°C.
Figure 8

Size distribution of the proteoliposomes containing wild-type and variants of AmtB.
Dynamic light scattering was used to determine the number-weighted distribution of liposome sizes in the detection reagent. The distribution was monodisperse, with a mean diameter of 110 nm.
To ensure that all AmtB variants were correctly inserted into the proteoliposomes, the proteoliposomes were solubilized in 2% DDM and the proteins analyzed by size exclusion chromatography using a superdex 200 (10 × 300) enhanced column. The elution profile of all variants and the wild-type were identical, showing a single monodisperse peak eluting between 10.4–10.6 ml (Figure 4—figure supplement 1). This demonstrated that all proteins were correctly folded, as trimers, in the proteoliposomes.
Solid supported membrane electrophysiology
Request a detailed protocolTo form the solid-supported membrane, 3 mm gold-plated sensors were prepared according to the manufacturer’s instructions (Nanion Technologies, Munich, Germany), as described previously (Bazzone et al., 2017). Proteoliposomes/empty liposomes were defrosted and sonicated in a sonication bath at 35 W for 1 min, diluted 10 times in non-activating (NA) solution (Supplementary file 1), and then 10 µl were added at the surface of the SSM on the sensor. After centrifugation, the sensors were stored at 4°C for a maximum of 48 hr before electrophysiological measurements. For the D2O experiments, all the solutions were prepared using D2O instead of water.
All measurements were made at room temperature (21°C) using a SURFE2R N1 apparatus (Nanion Technologies, Munich, Germany) with default parameters (Bazzone et al., 2017). Prior to any measurements, the quality of the sensors was determined by measuring their capacitance (15–30 nF) and conductance (<5 nS).
For functional measurements at a fixed pH, a single solution exchange protocol was used with each phase lasting 1 s (Bazzone et al., 2017). First, non-active (NA) solution was injected onto the sensor, followed by activating (A) solution containing the substrate at the desired concentration and finally NA solution (Supplementary file 1).
For the measurements under inwardly orientated pH gradient, a double solution exchange protocol was used (Bazzone et al., 2017), in which an additional resting solution phase of 15 min in NA solution at pH 8 was added to the end. The incubation phase adjusts the inner pH of the proteoliposomes to pH 8 and establishes a pH gradient at the beginning of each measurement by pulsing the activation solution at pH 5.
Each sensor was measured in the order pH (in/out) 7/7, 8/5 (with NH4+), 8/5 (without NH4+), and finally again 7/7 to ensure that the signals do not significantly decrease with time. The data are normalized against the measurements conducted at pH7 in/out for each sensor. All measurements were recorded on 8 sensors from two independent protein purification batches, with 3 measurements recorded for each sensor.
The kinetic parameters were calculated using Graphpad Prism 6 (GraphPad Software, San Diego, California, USA) and fitted according to the Michaelis-Menten equation (Key resources table). Lifetime analysis of the current (decay time of the transient current) was performed to differentiate small pre-steady state currents, which arise due to the binding of a charged species to membrane proteins, from currents reflecting full transport cycles, which show faster decay rates under raised liposomal LPR (Bazzone et al., 2017). The decay time of the transient current (Table 1) was calculated by fitting the raw transient current data between the apex of the peak and the baseline (after transport) with a non-linear regression using OriginPro 2017 (OriginLab, Northampton, Massachusetts, USA). The regression was done using a one-phase exponential decay function with time constant parameter:
The fit was obtained using the Levenberg-Marquardt iteration algorithm, where x and y represent coordinates on the respective axis, y0 represents the offset at a given point, A represents the amplitude, and t is the time constant.
Molecular Dynamics simulations
Request a detailed protocolThe AmtB trimer (PDB code: 1U7G) (Khademi et al., 2004) was processed using the CHARMM-GUI web server (Lee et al., 2016). Any mutations inserted during the crystallization process were reverted to the wild-type form. The N-termini and C-termini of the subunits were capped with acetyl and N-methyl amide moieties, respectively. The protein was then inserted into a membrane patch of xy-dimensions 13 × 13 nm. Unless otherwise specified, a membrane composition of palmitoyl oleoyl phosphatidyl ethanolamine and palmitoyl oleoyl phosphatidyl glycine (POPE/POPG) at a 3:1 ratio was used in order to approximate the composition of a bacterial cytoplasmic membrane. We employed the CHARMM36 forcefield for the protein and counter ions (Best et al., 2012). The water molecules were modeled with the TIP3P model (Jorgensen et al., 1983). Water bonds and distances were constrained by the Settle method (Miyamoto and Kollman, 1992), and all other bonds by the LINCS method (Hess et al., 1997). In simulations without ammonium, K+ and Cl- ions were added to neutralize the system and obtain a bulk ionic concentration of 250 mM. In simulations with ammonium, K+ was replaced by NH4+. The parameters for NH4+ and NH3 (umbrella sampling simulations) were adapted from Nygaard et al., 2006.
After a steepest descent energy minimization, the system was equilibrated by six consecutive equilibration steps using position restraints on heavy atoms of 1000 kJ/mol.nm2. The first three equilibration steps were conducted in an NVT ensemble, applying a Berendsen thermostat (Berendsen et al., 1984) to keep the temperature at 310K. The subsequent steps were conducted under an NPT ensemble, using a Berendsen barostat (Berendsen et al., 1984) to keep the pressure at 1 bar. Production molecular dynamics simulations were carried out using a v-rescale thermostat (Bussi et al., 2007) with a time constant of 0.2 ps, and a Berendsen barostat with semi-isotropic coupling. A timestep of 2 fs was used throughout the simulations.
In a subset of simulations, we aimed to test the effect of membrane voltage on the internal hydration of AmtB using CompEL. For the CompEL simulations (Kutzner et al., 2016), the system was duplicated along the z-axis, perpendicular to the membrane surface. To focus on the physiologically relevant voltage gradient in E. coli, that is a negative potential on the inside of the cell of magnitude −140 to −170 mV (Cohen and Venkatachalam, 2014), an antiparallel orientation of the two trimers in the double bilayers was used (Felle et al., 1980). The final double system consisted of a rectangular box of 13 × 13×20 nm. For the CompEL simulations, 1000 positively charged (either NH4+ or K+) and 1000 negatively charged ions (Cl-) were added to the system, then the system was neutralized, and the desired ion imbalance established.
The Umbrella Sampling (US) Potential-of-Mean-Force (PMF) calculations (Torrie and Valleau, 1977) were set up as described previously by Hub et al., 2010b. A snapshot was taken from the simulation of the single bilayer system with the twin-His motif in the DE protonation state and both the CWW and PWW occupied. The pore coordinates were obtained using the software HOLE (Smart et al., 1996), removing the solvent and mutating F215 to alanine during the HOLE run only. Starting coordinates for each umbrella window were generated by placing NH3 in the central x-y coordinate of the pore defined by HOLE at positions every 0.5 Å in the z coordinate. Solvent molecules within 2 Å of the ammonia's N atom were removed. Minimization and equilibration were then re-performed as described above. Unless otherwise stated, position restraints were used for all water oxygen atoms in the CWW with a 200 kJ/mol.nm2 force constant; while the TIP3 molecules within the lower water wire were not restrained. For the US the N atom of ammonia was position-restrained with a force constant of 1000 kJ/mol.nm2 on the z axis and a 400 kJ/mol.nm2 cylindrical flat-bottomed potential with a radius of 5 Å in the x-y plane, as described earlier by Hub et al., 2010a. For some US window simulations, the ammonia z-axis restraints were increased and the time step reduced in the equilibration to relax steric clashes between sidechains and ammonia. After equilibration, US simulations were run for 10ns, using the parameters described above (Lee et al., 2016) and removing the initial two ns for further equilibration. The PMF profiles were generated with the GROMACS implementation of the weighted histogram analysis method (WHAM) with the periodic implementation (Hub et al., 2010a). Further US simulations were performed to as needed to improve sampling in regions of the profile that were not sufficiently sampled. The Bayesian bootstrap method was performed with 200 runs to calculate the standard deviation of the PMF.
Free energy calculations for proton translocation
Request a detailed protocolThe free energies for proton translocation were evaluated by protonating the water molecules at different sites along the periplasmic and cytoplasmic water wires. Electrostatic effects in proteins are often treated more effectively using semi-macroscopic models which can overcome the convergence problems of more rigorous microscopic models. Here we used the semi-macroscopic protein dipole/Langevin dipole approach of Warshel and coworkers in the linear response approximation version (PDLD/S-LRA) (Kato et al., 2006; Sham et al., 2000). Positions of the water molecules in the PWW and CWW were obtained from the corresponding MD snapshots (Figure 1). All PDLD/S-LRA pKa calculations were performed using the automated procedure in the MOLARIS simulations package (Lee et al., 1993) in combination with the ENZYMIX force field. The simulation included the use of the surface‐constrained all atom solvent model (SCAAS) (Warshel and King, 1985) and the local reaction field (LRF) long-range treatment of electrostatics. At each site, 20 configurations for the charged and uncharged state were generated. The obtained pKa values were then converted to free energies for proton translocation.
Data availability
All data generated or analysed during this study are included in the manuscript and supporting files. Source data files have been provided for Figures 1-5 and Table 2. Simulation code is available on GitHub at https://github.com/UZgroup/A-two-lane-mechanism-for-selective-biological-ammonium-transport/ (copy archived at https://github.com/elifesciences-publications/A-two-lane-mechanism-for-selective-biological-ammonium-transport) and the trajectory files are available on Figshare (https://doi.org/10.6084/m9.figshare.12826316).
-
figshareMolecular dynamics simulation trajectories, AmtB in twin-His HSD-HSE and HSE-HSD states.https://doi.org/10.6084/m9.figshare.12826316
References
-
Similarities and differences between potassium and ammonium ions in liquid water: a first-principles studyPhysical Chemistry Chemical Physics 22:2540–2548.https://doi.org/10.1039/C9CP06163K
-
SSM-Based electrophysiology for transporter researchMethods in Enzymology 594:31–83.https://doi.org/10.1016/bs.mie.2017.05.008
-
Molecular dynamics with coupling to an external bathThe Journal of Chemical Physics 81:3684–3690.https://doi.org/10.1063/1.448118
-
Optimization of the additive CHARMM all-atom protein force field targeting improved sampling of the backbone φ, ψ and side-chain χ(1) and χ(2) dihedral anglesJournal of Chemical Theory and Computation 8:3257–3273.https://doi.org/10.1021/ct300400x
-
Canonical sampling through velocity rescalingThe Journal of Chemical Physics 126:014101.https://doi.org/10.1063/1.2408420
-
Bringing bioelectricity to lightAnnual Review of Biophysics 43:211–232.https://doi.org/10.1146/annurev-biophys-051013-022717
-
Merging In-Solution X-ray and neutron scattering data allows fine structural analysis of Membrane-Protein detergent complexesThe Journal of Physical Chemistry Letters 9:3910–3914.https://doi.org/10.1021/acs.jpclett.8b01598
-
Media preparation and bacteriological toolsCurrent Protocols in Molecular Biology 59:1.https://doi.org/10.1002/0471142727.mb0101s59
-
Quantitative measurements of membrane potential in Escherichia coliBiochemistry 19:3585–3590.https://doi.org/10.1021/bi00556a026
-
A twin histidine motif is the core structure for high-affinity substrate selection in plant ammonium transportersJournal of Biological Chemistry 295:3362–3370.https://doi.org/10.1074/jbc.RA119.010891
-
Improved method for high efficiency transformation of intact yeast cellsNucleic Acids Research 20:1425.https://doi.org/10.1093/nar/20.6.1425
-
The molecular basis of K+ exclusion by the Escherichia coli ammonium channel AmtBThe Journal of Biological Chemistry 288:14080–14086.https://doi.org/10.1074/jbc.M113.457952
-
LINCS: a linear constraint solver for molecular simulationsJournal of Computational Chemistry 18:1463–1472.https://doi.org/10.1002/(SICI)1096-987X(199709)18:12<1463::AID-JCC4>3.0.CO;2-H
-
Competition between uptake of ammonium and potassium in barley and Arabidopsis roots: molecular mechanisms and physiological consequencesJournal of Experimental Botany 61:2303–2315.https://doi.org/10.1093/jxb/erq057
-
The Rh protein family: gene evolution, membrane biology, and disease associationCellular and Molecular Life Sciences 67:1203–1218.https://doi.org/10.1007/s00018-009-0217-x
-
g_wham—A Free Weighted Histogram Analysis Implementation Including Robust Error and Autocorrelation EstimatesJournal of Chemical Theory and Computation 6:3713–3720.https://doi.org/10.1021/ct100494z
-
Potentials of mean force and permeabilities for carbon dioxide, Ammonia, and water flux across a rhesus protein channel and lipid membranesJournal of the American Chemical Society 132:13251–13263.https://doi.org/10.1021/ja102133x
-
Protonation states of Ammonia/ammonium in the hydrophobic pore of Ammonia transporter protein AmtBJournal of the American Chemical Society 129:1210–1215.https://doi.org/10.1021/ja066208n
-
A cis-dominant regulatory mutation linked to the argB-argC gene cluster in Saccharomyces cerevisiaeJournal of Molecular Biology 139:691–704.https://doi.org/10.1016/0022-2836(80)90055-8
-
Ammonium sensing in Escherichia coli. Role of the ammonium transporter AmtB and AmtB-GlnK complex formationThe Journal of Biological Chemistry 279:8530–8538.https://doi.org/10.1074/jbc.M312399200
-
An unusual twin-his arrangement in the pore of Ammonia channels is essential for substrate conductanceJournal of Biological Chemistry 281:39492–39498.https://doi.org/10.1074/jbc.M608325200
-
Comparison of simple potential functions for simulating liquid waterThe Journal of Chemical Physics 79:926–935.https://doi.org/10.1063/1.445869
-
The barrier for proton transport in aquaporins as a challenge for electrostatic models: The role of protein relaxation in mutational calculationsProteins: Structure, Function, and Bioinformatics 64:829–844.https://doi.org/10.1002/prot.21012
-
Insights into the function of ion channels by computational electrophysiology simulationsBiochimica Et Biophysica Acta (BBA) - Biomembranes 1858:1741–1752.https://doi.org/10.1016/j.bbamem.2016.02.006
-
A stable water chain in the hydrophobic pore of the AmtB ammonium transporterBiophysical Journal 92:L82–L84.https://doi.org/10.1529/biophysj.106.102756
-
Microscopic and semimicroscopic calculations of electrostatic energies in proteins by the POLARIS and ENZYMIX programsJournal of Computational Chemistry 14:161–185.https://doi.org/10.1002/jcc.540140205
-
CHARMM-GUI input generator for NAMD, GROMACS, AMBER, OpenMM, and CHARMM/OpenMM simulations using the CHARMM36 additive force fieldJournal of Chemical Theory and Computation 12:405–413.https://doi.org/10.1021/acs.jctc.5b00935
-
Uniport of NH4+ by the root hair plasma membrane ammonium transporter LeAMT1;1Journal of Biological Chemistry 277:13548–13555.https://doi.org/10.1074/jbc.M200739200
-
A family of ammonium transporters in Saccharomyces cerevisiaeMolecular and Cellular Biology 17:4282–4293.https://doi.org/10.1128/MCB.17.8.4282
-
Ammonium ion transport by the AMT/Rh homologue LeAMT1;1Biochemical Journal 396:431–437.https://doi.org/10.1042/BJ20060051
-
Evolution of electrogenic ammonium transporters (AMTs)Frontiers in Plant Science 7:352.https://doi.org/10.3389/fpls.2016.00352
-
Settle: An analytical version of the SHAKE and RATTLE algorithm for rigid water modelsJournal of Computational Chemistry 13:952–962.https://doi.org/10.1002/jcc.540130805
-
Ammonium recruitment and Ammonia transport by E. coli Ammonia channel AmtBBiophysical Journal 91:4401–4412.https://doi.org/10.1529/biophysj.106.089714
-
Examining methods for calculations of binding free energies: lra, LIE, PDLD-LRA, and PDLD/S-LRA calculations of ligands binding to an HIV proteaseProteins: Structure, Function, and Genetics 39:393–407.https://doi.org/10.1002/(SICI)1097-0134(20000601)39:4<393::AID-PROT120>3.0.CO;2-H
-
HOLE: A program for the analysis of the pore dimensions of ion channel structural modelsJournal of Molecular Graphics 14:354–360.https://doi.org/10.1016/S0263-7855(97)00009-X
-
Membrane topology of the Mep/Amt family of ammonium transportersMolecular Microbiology 37:331–344.https://doi.org/10.1046/j.1365-2958.2000.01994.x
-
Nonphysical sampling distributions in Monte Carlo free-energy estimation: Umbrella samplingJournal of Computational Physics 23:187–199.https://doi.org/10.1016/0021-9991(77)90121-8
-
Structural basis for Mep2 ammonium transceptor activation by phosphorylationNature Communications 7:11337.https://doi.org/10.1038/ncomms11337
-
Ammonium transporters achieve charge transfer by fragmenting their substrateJournal of the American Chemical Society 134:10419–10427.https://doi.org/10.1021/ja300129x
-
Functional and physiological evidence for a rhesus-type Ammonia transporter in Nitrosomonas europaeaFEMS Microbiology Letters 273:260–267.https://doi.org/10.1111/j.1574-6968.2007.00805.x
Article and author information
Author details
Mélanie Boeckstaens

Gaëtan Dias Mirandela
Funding
Tenovus (S17-07)
- Arnaud Javelle
Scottish Universities Physics Alliance
- Ulrich Zachariae
Natural Environment Research Council (NE/M001415/1)
- Paul A Hoskisson
Fonds De La Recherche Scientifique - FNRS (WELBIO grant ref: CR-2019A-05R.)
- Anna-Maria Marini
The funders had no role in study design, data collection and interpretation, or the decision to submit the work for publication.
Acknowledgements
Special thanks to Prof. Iain Hunter (Strathclyde Institute of Pharmacy and Biomedical Sciences) for invaluable discussions and help during this project. We also thank Pascale Van Vooren for technical support and Thomas P Jahn for sharing the triple-mepΔ trk,1,2Δ yeast strain. Anna Maria Marini is a senior research associate FNRS and WELBIO investigator and thanks the help of FRFS-WELBIO grant ref: CR-2019A-05R.
Copyright
© 2020, Williamson et al.
This article is distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use and redistribution provided that the original author and source are credited.
Metrics
-
- 3,016
- views
-
- 389
- downloads
-
- 25
- citations
Views, downloads and citations are aggregated across all versions of this paper published by eLife.
Download links
A two-part list of links to download the article, or parts of the article, in various formats.
Downloads (link to download the article as PDF)
Open citations (links to open the citations from this article in various online reference manager services)
Cite this article (links to download the citations from this article in formats compatible with various reference manager tools)
A two-lane mechanism for selective biological ammonium transport
eLife 9:e57183.
https://doi.org/10.7554/eLife.57183
Further reading
-
- Biochemistry and Chemical Biology
- Computational and Systems Biology
The spike protein is essential to the SARS-CoV-2 virus life cycle, facilitating virus entry and mediating viral-host membrane fusion. The spike contains a fatty acid (FA) binding site between every two neighbouring receptor-binding domains. This site is coupled to key regions in the protein, but the impact of glycans on these allosteric effects has not been investigated. Using dynamical nonequilibrium molecular dynamics (D-NEMD) simulations, we explore the allosteric effects of the FA site in the fully glycosylated spike of the SARS-CoV-2 ancestral variant. Our results identify the allosteric networks connecting the FA site to functionally important regions in the protein, including the receptor-binding motif, an antigenic supersite in the N-terminal domain, the fusion peptide region, and another allosteric site known to bind heme and biliverdin. The networks identified here highlight the complexity of the allosteric modulation in this protein and reveal a striking and unexpected link between different allosteric sites. Comparison of the FA site connections from D-NEMD in the glycosylated and non-glycosylated spike revealed that glycans do not qualitatively change the internal allosteric pathways but can facilitate the transmission of the structural changes within and between subunits.




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}