Embryonic origins of forebrain oligodendrocytes revisited by combinatorial genetic fate mapping
- Institutes of Brain Science, State Key Laboratory of Medical Neurobiology and MOE Frontiers Center for Brain Science, Department of Neurobiology, Zhongshan Hospital, Fudan University, China
eLife assessment
In this study the authors revisited the question of the embryonic origin of telencephalic oligodendrocytes using some new and powerful genetic tools. There is convincing evidence to support previous suggestions of a predominantly cortical origin of oligodendrocytes in the cerebral cortex, however the new studies suggest that LGE/CGE-derived oligodendrocytes make a modest contribution in some areas, while MGE/POA-derived oligodendrocytes make a small but enduring contribution. The findings are valuable and should be of interest to developmental and myelin biologists.
https://doi.org/10.7554/eLife.95406.3.sa0Significance of the findings:
Valuable: Findings that have theoretical or practical implications for a subfield
- Landmark
- Fundamental
- Important
- Valuable
- Useful
Strength of evidence:
Convincing: Appropriate and validated methodology in line with current state-of-the-art
- Exceptional
- Compelling
- Convincing
- Solid
- Incomplete
- Inadequate
During the peer-review process the editor and reviewers write an eLife Assessment that summarises the significance of the findings reported in the article (on a scale ranging from landmark to useful) and the strength of the evidence (on a scale ranging from exceptional to inadequate). Learn more about eLife Assessments
Abstract
Multiple embryonic origins give rise to forebrain oligodendrocytes (OLs), yet controversies and uncertainty exist regarding their differential contributions. We established intersectional and subtractional strategies to genetically fate map OLs produced by medial ganglionic eminence/preoptic area (MGE/POA), lateral/caudal ganglionic eminences (LGE/CGE), and dorsal pallium in the mouse brain. We found that, contrary to the canonical view, LGE/CGE-derived OLs make minimum contributions to the neocortex and corpus callosum, but dominate piriform cortex and anterior commissure. Additionally, MGE/POA-derived OLs, instead of being entirely eliminated, make small but sustained contribution to cortex with a distribution pattern distinctive from those derived from the dorsal origin. Our study provides a revised and more comprehensive view of cortical and white matter OL origins, and established valuable new tools and strategies for future OL studies.
Introduction
Oligodendrocytes (OLs) are an important class of macroglia responsible for producing the myelin sheaths that insulate and protect neuronal axons. Forebrain OLs arise from multiple embryonic origins. Previous fate-mapping study using Nkx2.1Cre (Xu et al., 2008), Gsh2Cre (Kessaris et al., 2006), and Emx1Cre (Gorski et al., 2002) reported consecutive and competing waves of OLs derived from medial ganglionic eminence/preoptic area (MGE/POA), lateral/caudal ganglionic eminences (LGE/CGE), and dorsal pallium (Kessaris et al., 2006). The first wave of OLs generated by MGE/POA (MPOLs) was believed to be eliminated postnatally, while those from the second and third waves (LCOLs and dOLs) survive and populate the cortex and corpus callosum at comparable proportions. Several other studies provided both supporting and contradicting evidence to this model (Nakahira et al., 2006; Tsoa et al., 2014; Naruse et al., 2016; Orduz et al., 2019; Liu et al., 2021; Shen et al., 2021; Tripathi et al., 2011; Winkler et al., 2018). Moreover, Gsh2 was recently found to be expressed in dorsal progenitors (Zhang et al., 2020), casting doubt on the interpretation of lineage tracing data from Gsh2Cre.
In this study, we generated new genetic tools and combinatorial fate-mapping strategies which allow direct visualization and comparison among OLs derived from different origins. We found that neocortical OLs are primarily composed of dOLs, rather than similar proportions of LCOLs and dOLs. In contrast, LCOLs and dOLs made comparable contributions to piriform cortex. We also found that although MPOLs only make a small contribution, they do persist in the cortex beyond adulthood with a unique spatial pattern distinct from that of the dOLs. In the two major white matter commissure tracts, dOLs are the vast majority in corpus callosum but make little contribution to anterior commissure, while LCOLs behaved the opposite. These findings significantly revised the classical view and provided a new and more comprehensive picture of cortical and white matter OL origins.
Results and discussion
To unambiguously track OLs from different embryonic origins, we first generated a knock-in driver, OpalinP2A-Flpo-T2A-tTA2 (Figure 1), orthogonal to Cre drivers that label dorsal or ventral progenitors (ProgenitorCre). Opalin (also known as Tmem10) encodes oligodendrocytic myelin paranodal and inner loop protein that are specifically expressed in differentiated OLs (Kippert et al., 2008; Yoshikawa et al., 2008; Jiang et al., 2013; Marques et al., 2016). In OpalinP2A-Flpo-T2A-tTA2, Flpo and tTA2 were inserted before the STOP codon and linked by self-cleavage peptide P2A and T2A (Figure 1A–C), allowing co-transcription and translation with Opalin. Flp-mediated recombination by this driver (hereinafter referred to as OpalinFlp for simplicity) enables highly specific, efficient, and irreversible OL labeling, while the tTA2 component offers the flexibility for OL-specific labeling in tunable densities (Figure 1D–F).
Figure 1
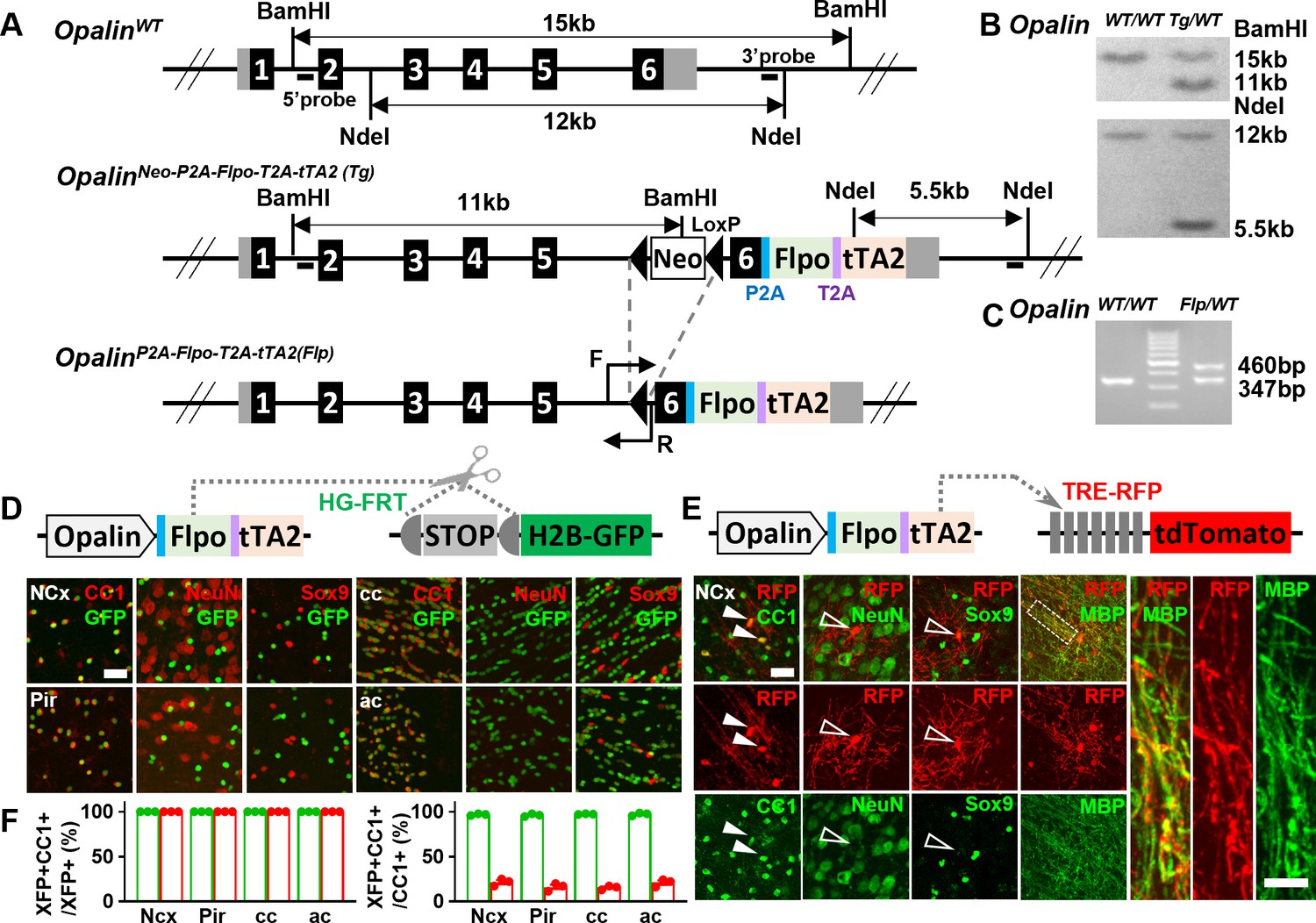
A new driver mouse for efficient and specific oligodendrocyte (OL) labeling.
(A) Scheme for generating the OpalinP2A-Flpo-T2A-tTA2 allele. (B) Southern blot confirmation of correctly targeted embryonic stem cell clone. (C) Genomic polymerase chain reaction (PCR) to genotype F1 offspring. (D) OL labeling by Flp. (E) OL labeling by tTA2. High magnification images of the boxed region showing co-localization of red fluorescent protein (RFP) with myelin basic protein (MBP) staining, which further demonstrated the myelination ability of labeled OLs. (F) Quantification of labeling specificity (left panel) and efficiency (right panel) by colacalization with OL marker CC1. Both reporting systems are highly specific, as shown by the complete co-localization of fluorescent protein (XFP) with OL marker (CC1) and lack of co-staining with neuronal marker (NeuN) or astrocyte marker (Sox9). Quantification bar graph was not presented for NeuN and Sox9 as zero co-localizations were observed in all analyzed regions. Close to complete OL labeling was achieved by Flp-dependent H2B-GFP reporter in all analyzed regions (green dots), while sparser labeling with variable regional density was achieved by tTA2-dependent tdTomato reporter driven by TRE promoter (red dots). NCx: neocortex. Pir: piriform cortex. cc: corpus callosum. ac: anterior commissure. Scale bar: 50 μm in low magnification images, 5 μm in high magnification images. Quantification: n = 3. Dots represent data from individual mice.
-
Figure 1—source data 1
Raw unedited blot for Figure 1B.
- https://cdn.elifesciences.org/articles/95406/elife-95406-fig1-data1-v1.zip
-
Figure 1—source data 2
Uncropped and labeled blot for Figure 1B.
- https://cdn.elifesciences.org/articles/95406/elife-95406-fig1-data2-v1.zip
-
Figure 1—source data 3
Raw unedited gel for Figure 1C.
- https://cdn.elifesciences.org/articles/95406/elife-95406-fig1-data3-v1.zip
-
Figure 1—source data 4
Uncropped and labeled gel for Figure 1C.
- https://cdn.elifesciences.org/articles/95406/elife-95406-fig1-data4-v1.zip
-
Figure 1—source data 5
The raw data for the visualization of data presented in Figure 1F.
- https://cdn.elifesciences.org/articles/95406/elife-95406-fig1-data5-v1.xlsx
Next, we established two types of genetic combinatorial fate-mapping strategies to directly visualize OLs from different embryonic origins (Figure 2): (1) combining OpalinFlp and ProgenitorCre with intersectional reporters Ai65 to label OLs derived from Cre+ progenitor domain by RFP (Figure 2A); (2) combining OpalinFlp and ProgenitorCre with RC::FLTG (Plummer et al., 2015) to simultaneously label OLs derived from Cre+ progenitors by green fluorescent protein (GFP) and OLs derived from the complementing Cre− progenitors by RFP (Figure 2B, Figure 2—figure supplement 1A). The first approach allowed us to track dOLs and MPOLs (Figure 2C, E). The second approach empowered us to observe and compare OLs generated from dorsal and ventral origins (Figure 2—figure supplement 1B), or those from Gsh2+ and Gsh2− progenitors, in the same brain (Figure 2—figure supplement 1C). Importantly, the subtraction power enabled us to target OLs derived from LGE/CGE progenitors that express neither Emx1 nor Nkx2.1 (Figure 2D and Figure 2—figure supplement 1D). In addition, these strategies greatly facilitated the identification of OLs derived from specific origin which exist at relatively low density in certain regions.
Figure 2 with 3 supplements see all
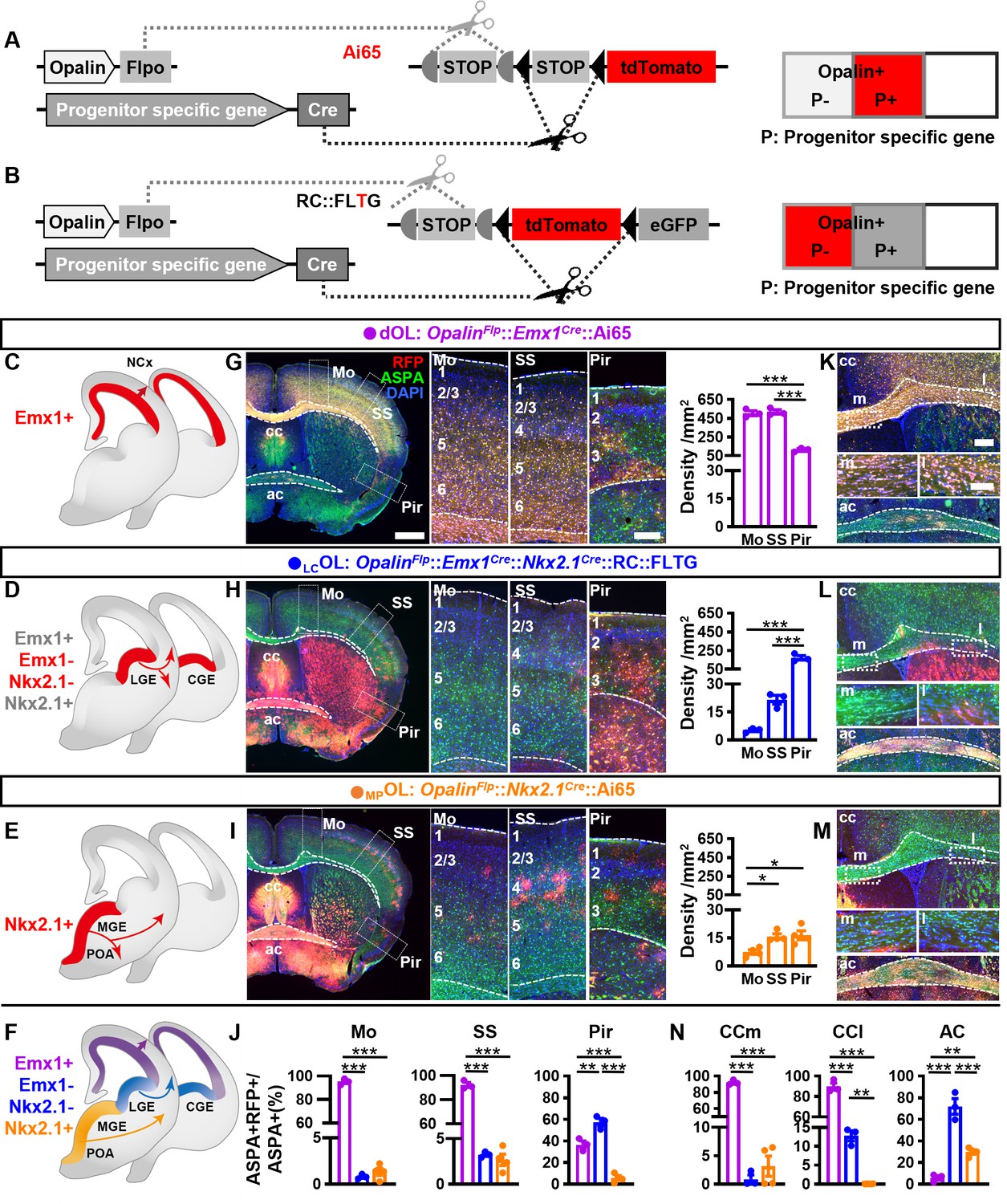
Combinatorial fate mapping of dOLs, MPOLs, and LCOLs.
(A) Strategy for intersectional labeling. Flp-AND-Cre labels oligodendrocytes (OLs) from Cre-expressing progenitors with RFP. (B) Strategy for subtractional labeling of OLs derived from non-Cre-expressing progenitors with RFP. The eGFP expressing OLs derived from Cre-expressing progenitors were not used for analysis in this scenario and thereby were not highlighted by color. Schematics showing intersectional labeling of dOLs in OpalinFlp::Emx1Cre::Ai65 (C), subtractional labeling of LCOLs in OpalinFlp::Emx1Cre::Nkx2.1Cre::RC::FLTG (D), intersectional labeling of MPOLs in OpalinFlp::Nkx2.1Cre::Ai65 (E), and cortical OLs derived from all three origins (F). (G–I) Representative images (left panels) and quantifications (right panels) of RFP+ cell density in motor cortex (Mo), somatosensory cortex (SS), and piriform cortex (Pir). (J) Quantification of differential contribution to ASPA+ OLs by three embryonic origins to Mo, SS, and Pir. Representative images (K–M) and quantifications (N) of differential contribution to ASPA+ OLs by three embryonic origins in the two major commissure white matter tracts: corpus callosum (cc) and anterior commissure (ac). MPOLs and LCOLs preferentially reside in the medial and lateral cc (cc-m and cc-l), respectively. Scale bar: 1 mm in low magnification images in (G–I), 250 μm in high magnification images of the boxed area in (G–I) and low magnification images in (K–M), 100 μm in high magnification images of the boxed area (cc-m and cc-l) in (K–M). n = 3 for dOLs and LCOLs; n = 4 for MPOLs. Dots represent data from individual mice. Error bar: standard error of the mean (SEM). *p < 0.05, **p < 0.01, ***p < 0.001.
-
Figure 2—source data 1
The raw data for the visualization of data presented in Figure 2G–J, N.
- https://cdn.elifesciences.org/articles/95406/elife-95406-fig2-data1-v1.xlsx
Deploying these strategies, we assessed the differential contributions of dOLs, LCOLs, and MPOLs by analyzing RFP+ cells in the following mice: OpalinFlp::Emx1Cre::Ai65 (Figure 2C), OpalinFlp::Emx1Cre::Nkx2.1Cre::RC::FLTG (Figure 2D), and OpalinFlp::Nkx2.1Cre::Ai65 (Figure 2E). To better assess their contributions to the total OL population (Figure 2F), we co-stained RFP with the mature OL marker aspartoacylase (ASPA) (Huang et al., 2023; Figure 2G–I) and quantified the ratio of co-localization (Figure 2J). Notably, all RFP+ cells are ASPA+, reassured the specificity of our label strategies. We observed two significant differences from the traditional model in the neocortex. The first major deviation is that, instead of comparable contributions by dOLs and LCOLs, the vast majority of neocortical OLs were dOLs but not LCOLs. The densities (Figure 2G–I and Figure 2—figure supplement 2A–F) and ASPA ratios (Figure 2J) of dOLs are much higher than those of LCOLs. Considering the possibility of incomplete recombination in combinatorial reporters, and the relatively low Cre activity in the dorsal MGE of Nkx2.1Cre (Xu et al., 2008), the genuine contribution of LCOLs to the neocortex could be even lesser than our current observation. Therefore, the large quantity of neocortical OLs labeled by Gsh2Cre in previous study (Kessaris et al., 2006) or by GFP in OpalinFlp::Gsh2Cre::RC::FLTG (Figure 2—figure supplement 1C) most likely were predominantly dOLs generated by Gsh2+ dorsal progenitors (Zhang et al., 2020), rather than bona fide LCOLs.
The second major deviation is that cortical MPOLs are not completely depleted postnatally. Instead, they make a small but continued contribution with a unique spatial distribution pattern (Figure 2I, J and Figure 2—figure supplement 2D–G). MPOLs display a clear rostrocaudal density decline (Figure 2—figure supplement 2E, F), a higher density in somatosensory cortex (SS) than motor cortex (Mo) (Figure 2I), and a laminar preference toward layer 4 (L4) in SS (Figure 2—figure supplement 2G). In contrast, the distribution of dOLs and LCOLs do not vary significantly across the rostrocaudal axis (Figure 2—figure supplement 2E, F) or between Mo and SS (Figure 2G, H), but exhibits increased density toward deeper layers (Figure 2—figure supplement 2G). Importantly, we have observed cortical MPOLs in mice as old as 1 year (Figure 2—figure supplement 2H), well beyond the age analyzed in previous reports (Kessaris et al., 2006; Orduz et al., 2019; Liu et al., 2021), suggesting a persisted contribution.
We then turned our attention to the lateral three-layer archicortex, piriform cortex (Pir). Different from the neocortex, Pir contains higher proportions (Figure 2J) of LCOLs than dOLs. MPOLs make the lowest contribution (Figure 2J) at a density similar to SS and higher than Mo (Figure 2I).
These combinatorial models also grant us the opportunity to revisit the differential contributions of dOLs, LCOLs, and MPOLs to the two commissural white matter tracts, corpus callosum (cc), and anterior commissure (ac), which contain high density of OLs (Figure 2K–M). We found that, similar to the neocortex, cc is mainly populated by dOLs and supplemented by very low proportions of LCOLs and MPOLs (Figure 2K–N). Interestingly, LCOLs and MPOLs seem to show preferential distribution in the lateral and medial regions of cc (cc-l and cc-m), respectively (Figure 2L–N). Different from cc, ac is mainly populated by LCOLs and MPOLs and supplemented by very low proportion of dOLs (Figure 2K–N).
To substantiate the above results, we further breed OpalinFlp::Emx1Cre::Nkx2.1Cre::Ai65 to label dOLs together with MPOLs by RFP and co-stained them with ASPA (Figure 2—figure supplement 3). RFP−ASPA+ cells were difficult to find in Mo, SS, and cc, but were more easily observed in Pir and ac, consistent with the respective low and high LCOL contributions in these regions.
In summary, our findings significantly revised the canonical model of forebrain OL origins (Figure 3A), and provided a new and more comprehensive view (Figure 3B). We demonstrated that neocortical OLs are mainly derived from dorsal origin with small but lasting contribution from the ventral origin (Figure 2, Figure 2—figure supplements 1B and 2). Our data showed that LGE/CGE makes little contribution to neocortex and cc, but makes major contribution to piriform cortex and ac (Figure 2 and Figure 2—figure supplement 3). This finding is supported by another report in which in utero electroporation failed to label LGE-derived cortical OLs in both embryonic and early postnatal brains, and an exclusion strategy revealed very low percentage of LGE/CGE-derived cortical OLs in neonatal brains (Li et al., 2023). The lack of adult labeling in our study together with the lack of developmental labeling in the other study suggests that the lack of LCOL in neocortex is less likely caused by competitive postnatal elimination, but more likely due to limited production and/or allocation. We further discovered that MGE/POA makes a small but persistent contribution to the neocortex with a distinct distribution pattern featured by a rostral-high to caudal-low gradient and a preference toward L4 in SS (Figure 2—figure supplement 2). Whether their enduring existence and highly biased localization has functional implications awaits future exploration. In addition, we found that the cc showed a similar OL composition as the neocortex, but the Pir and the ac each exhibited distinct OL compositions in term of their embryonic origins. LCOLs are the major contributor to both regions, while dOLs and MPOLs mainly contribute to Pir and ac, respectively (Figure 2).
Figure 3
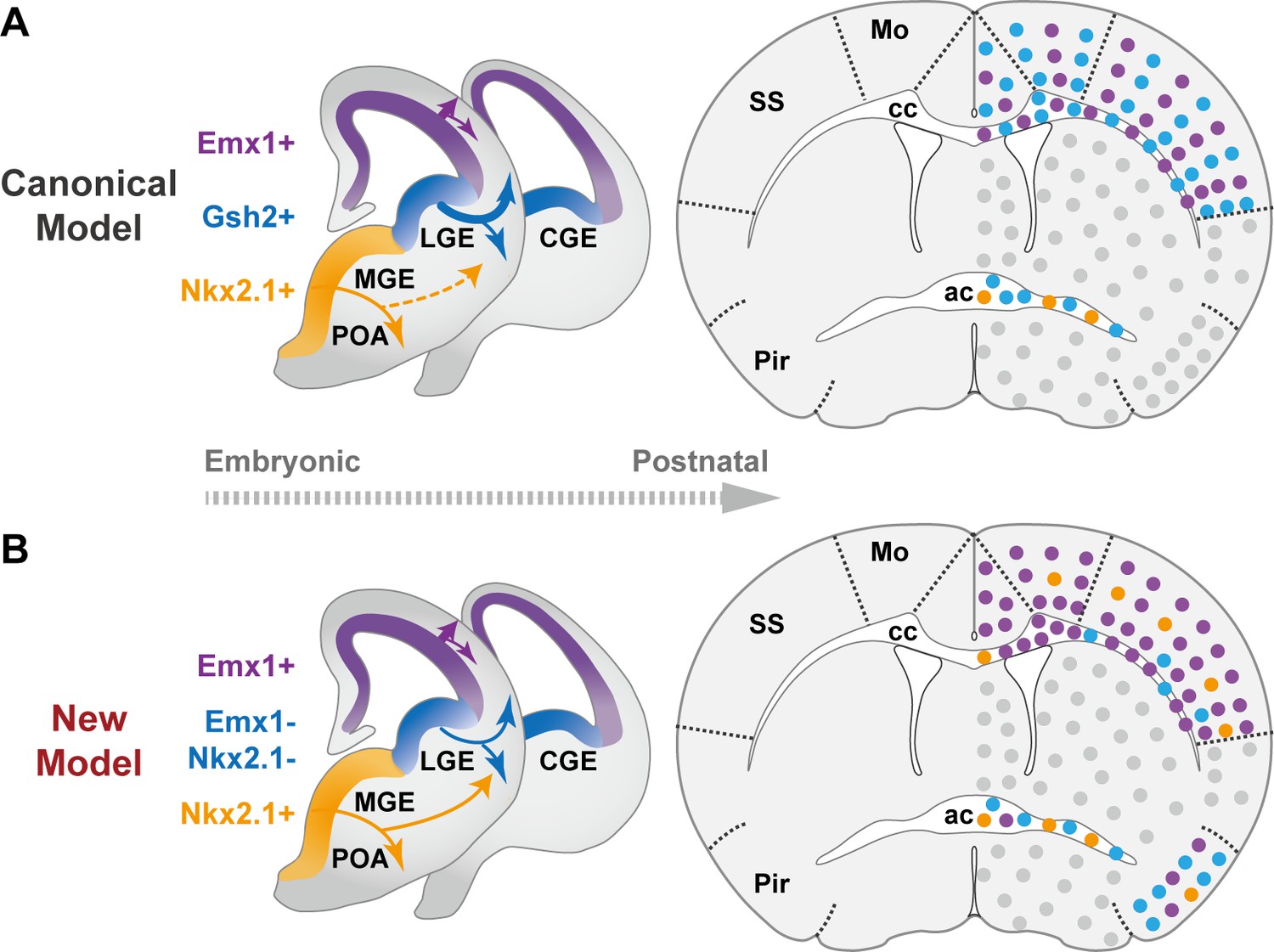
The classical and revised model of forebrain oligodendrocyte (OL) origins.
(A) In the classical model (Kessaris et al., 2006), OLs derived from medial ganglionic eminence/preoptic area (MGE/POA) (orange) were largely eliminated postnatally (thin dashed line), while those from lateral/caudal ganglionic eminences (LGE/CGE) (blue) and dorsal origin (purple) survive at similar proportions (thick solid line). Therefore, neocortex (NCx) and corpus callosum (cc) contain comparable density of LCOLs (blue dots) and dOLs (purple dots) and are devoid of MPOLs (orange dots). (B) In the new model, NCx and cc mainly contain dOLs with very low contribution from the ventral origins. LCOLs mainly contribute to piriform cortex (Pir) and anterior commissure (ac). MPOLs makes a small but sustained contribution to NCx, with a strong laminar preference toward layer 4 in somatosensory cortex (SS). In addition, dOLs and MPOLs also make substantial contributions to Pir and ac, respectively. Gray dots indicate OLs in unanalyzed regions.
In addition to the new framework of forebrain OL origins (Figure 3), we also generated a new driver (Figure 1) and established multiple combinatorial genetic models (Figure 2) for efficient tracking and direct visualization of OLs from different embryonic origins without interference from other cells types sharing the same progenitor domains such as OL precursors, astrocytes, and neurons (Figures 1–2). These tools set up a firm foundation and will provide reliable experimental access for future inquiries on the development and function of diverse OLs in healthy and disease brains (Gong et al., 2022), especially to uncover the relationship between their developmental origins and the functional and molecular heterogeneity.
Materials and methods
Key resources table
| Reagent type (species) or resource | Designation | Source or reference | Identifiers | Additional information |
|---|---|---|---|---|
| Genetic reagent (Mus musculus) | Nkx2.1Cre | The Jackson Laboratory | Strain#: 008661; RRID: IMSR_JAX:008661 | |
| Genetic reagent (Mus musculus) | Gsh2Cre | The Jackson Laboratory | Strain#: 025806; RRID: IMSR_JAX:025806 | |
| Genetic reagent (Mus musculus) | Emx1Cre | The Jackson Laboratory | Strain#: 005628; RRID: IMSR_JAX:005628 | |
| Genetic reagent (Mus musculus) | Ai65 | The Jackson Laboratory | Strain#: 021875; RRID: IMSR_JAX:021875 | |
| Genetic reagent (Mus musculus) | RC::FLTG | The Jackson Laboratory | Strain#: 026932; RRID: IMSR_JAX:026932 | |
| Genetic reagent (Mus musculus) | Ai62 | The Jackson Laboratory | Strain#: 022731; RRID: IMSR_JAX:022731 | |
| Genetic reagent (Mus musculus) | HG-FRT | The Jackson Laboratory | Strain#: 028581; RRID: IMSR_JAX:028581 | |
| Genetic reagent (Mus musculus) | OpalinP2A-Flpo-T2A-tTA2 | This paper | See Materials and methods, Mice | |
| Antibody | anti-RFP (goat polyclonal) | SICGEN | Cat# AB0081-200; RRID: AB_2333095 | IF (1:2000) |
| Antibody | anti-RFP (rabbit polyclonal) | Rockland | Cat# 600-401-379; RRID: AB_2209751 | IF (1:2000) |
| Antibody | anti-GFP (chicken polyclonal) | Aves Labs | Cat# GFP-1020; RRID: AB_10000240 | IF (1:1000) |
| Antibody | anti-MBP (rat polyclonal) | AbD Serotec | Cat# MCA409S; RRID: AB_325004 | IF (1:500) |
| Antibody | anti-CC1 (rabbit polyclonal) | Oasis Biofarm | Cat# OB-PRB070; RRID: AB_2934254 | IF (1:500) |
| Antibody | anti-CC1 (mouse polyclonal) | Millipore | Cat# OP80; RRID: AB_2057371 | IF (1:300) |
| Antibody | anti-ASPA (rat polyclonal) | Oasis Biofarm | Cat# OB-PRT005; RRID: AB_2938679 | IF (1:200) |
| Antibody | anti-Sox9 (rabbit polyclonal) | Chemicon | Cat# AB5535; RRID: AB_2239761 | IF (1:2000) |
| Antibody | anti-NeuN (mouse monoclonal) | Millipore | Cat# MAB377; RRID: AB_2298772 | IF (1:500) |
| Sequence-based reagent | Opalin-F | This paper | PCR primers | GGCCTATGTTTGATTTCCAGCACTG |
| Sequence-based reagent | Opalin-R | This paper | PCR primers | AGCACTTATGACTGCTGAGCCGTTC |
| Chemical compound, drug | Tail lysis buffer | Viagen | Cat# 102-T | |
| Chemical compound, drug | Proteinase K | Beyotime | Cat# ST535 | |
| Chemical compound, drug | Sodium pentobarbital | Sigma-Aldrich | Cat# P3761 | |
| Chemical compound, drug | Normal Donkey Serum | Abcam | Cat# ab7475 | |
| Chemical compound, drug | Triton X-100 | Sigma-Aldrich | Cat# X100PC | |
| Chemical compound, drug | Citrate buffer | Oasis-Biofarm | Cat# BR-AB001 | |
| Other | Aqua-mount | Southern Biotech | Cat# 0100-01 | |
| Other | DAPI stain | Invitrogen | Cat# D1306 | (10 mg/ml) |
| Software, algorithm | ImageJ | National Institutes of Health | RRID: SCR_003070 | |
| Software, algorithm | QuPath | Queen’s University Belfast | RRID: SCR_018257 | |
| Software, algorithm | Adobe Photoshop | Adobe Systems | RRID: SCR_014199 | |
| Software, algorithm | GraphPad Prism v8.0.1 | GraphPad Software | RRID: SCR_002798 |
Mice
All mouse studies were carried out in strict accordance with the guidelines of the Institutional Animal Care and Use Committee of School of Basic Medical Sciences, Fudan University. All husbandry and experimental procedures were reviewed and approved by the same committee (Permit Number: 20210302-137). All applicable institutional and/or national guidelines for the care and use of animals were followed. The following transgenic mouse lines were used in this study: Nkx2.1Cre (Jax 008661) (Xu et al., 2008), Gsh2Cre (Jax 025806) (Kessaris et al., 2006), Emx1Cre (Jax 005628) (Gorski et al., 2002), Ai65 (Jax 021875) (Madisen et al., 2015), and RC::FLTG (Jax 026932) (Plummer et al., 2015). The tTA2-dependent tdTomato reporter (TRE-RFP) was derived from Ai62 (Jax 022731) (Madisen et al., 2015), by removing LoxP-STOP-LoxP with E2a-Cre (Jax 003724). The Flp-dependent H2B-GFP reporter (HG-FRT) was derived from HG-dual (Jax 028581) via removal of loxP flanking STOP cassette by CMV-Cre (He et al., 2016). The OpalinP2A-Flpo-T2A-tTA2 allele was generated by targeted insertion of the T2A-Flpo-P2A-tTA2 sequence immediately before the STOP codon of the endogenous Opalin gene using homologous recombination. Gene targeting vector was generated using PCR-based cloning approach as described before (He et al., 2016). More specifically, a 4.7-kb 5′ homology arm, a loxP flanking Neo-positive selection cassette, a T2A-Flpo-P2A-tTA2 cassette and a 2.7-kb 3′ homology arm were cloned into a building vector containing the DTA-negative selection cassette to generate the targeting vector. Targeting vector was linearized and transfected into a C57/black6 ES cell line. ES clones that survived through negative and positive selections were first screened by genomic PCR, then confirmed by Southern blotting using appropriate DIG-dUTP-labeled probes. One positive ES cell clone was used for blastocyst injection to obtain male chimera mice carrying the modified allele following standard procedures. Chimera males were bred with C57BL/6J females to confirm germline transmission by genomic PCR. The Neo selection cassette was self-excised during spermatogenesis of F0 chimeras. Heterozygous F1 siblings were bred with one another to establish the colony. Targeting vector construction, ES cell transfections and screening, blastocyst injections, and chimera breeding were performed by Cyagen.
Genomic PCR
Request a detailed protocolGenomic DNA was prepared from mouse tails. Tissue was lysed by incubation in tail lysis buffer (Viagen, 102-T) with 0.1 mg/ml proteinase K (Diamond, A100706) overnight at 55°C followed by 45 min at 90°C in an air bath to inactivate proteinase K. The lysate was cleared by centrifugation at maximum speed (21,130 G) for 15 min in a table-top centrifuge. Supernatant containing genomic DNA was used as the PCR template for amplifying DNA products. The following primers were used:
Opalin-F: 5′-GGCCTATGTTTGATTTCCAGCACTG-3′
Opalin-R: 5′-AGCACTTATGACTGCTGAGCCGTTC-3′
Immunohistochemistry and microscopy
Request a detailed protocolMice were anesthetized by intraperitoneal injection of 1.5% sodium pentobarbital (0.09 mg/g body weight) and then intracardially perfused with saline followed by 4% paraformaldehyde in 0.1 M phosphate buffer. Following post fixation at 4°C for 24 hr, brain samples were sectioned at 30 μm using a vibratome (Leica VT1000S), or transferred into 30% sucrose in 0.1 M PB for cryoprotection, embedded in optimal cutting temperature (OCT) compound, and sectioned using a cryostat (Leica CM1950). For CC1 immunostaining, antigen retrieval was performed prior to blocking by boiling for 3 min in 10 mM citrate buffer (pH 6.0). Sections were blocked in phosphate buffered saline (PBS) containing 0.05% Triton and 5% normal donkey serum and then incubated with the following primary antibodies in the blocking solution at 4°C overnight: RFP (goat polyclonal antibody, 1:2000, SICGEN AB0081-200; rabbit polyclonal antibody, 1:2000, Rockland 600-401-379), GFP (chicken polyclonal antibody, 1:1000, Aves Labs, GFP-1020), MBP (rat polyclonal antibody, 1:500, AbD Serotec, MCA409S), CC1 (rabbit polyclonal antibody, 1:500, Oasis Biofarm, OB-PRB070, mouse polyclonal antibody, 1:300, Millipore, OP80), ASPA (rat polyclonal antibody, 1:200, Oasis Biofarm, OB-PRT005), Sox9 (rabbit polyclonal antibody, 1:2000, Chemicon, AB5535), and NeuN (mouse monoclonal antibody, 1:500, Millipore, MAB377). Sections were then incubated with appropriate Alexa fluor dye-conjugated IgG secondary antibodies (1:500, Thermo Fisher Scientific) or CF dye-conjugated IgG secondary antibodies (1:250, Sigma) in blocking solution and mounted in Aqua-mount (Southern Biotech, 0100-01). Sections were counterstained with DAPI (4',6-diamidino-2-phenylindole). Sections were imaged with confocal microscopy (Olympus FV3000), fluorescence microscopy (Nikon Eclipse Ni; Olympus VS120; Olympus VS200), and fluorescent stereoscope (Nikon SMZ25). All quantifications were performed in 2-month-old adult mice from coronal sections between Bregma +1.94 and −2.80 mm. Anatomical regions were identified according to the Paxinos ‘The Mouse Brain’ Atlas and the Allen Reference Atlas, and their areas were measured in ImageJ for density calculations, whenever applicable. For cortical regions, every fourth section within the range of selection was analyzed. For whiter matter tracts, three consecutive sections at Bregma 0.14 were analyzed. At least three brains were analyzed for each genotype. To quantify density and co-localization, cells were identified and counted in Adobe Photoshop or ImageJ in conjugation with QuPath.
Statistical analysis
Request a detailed protocolGraphPad Prism version 8.0.1 was used for statistical calculations. No statistical methods were used to predetermine sample sizes, but our sample sizes are similar to those reported in previous publications. Data collection and analysis were performed blind to the conditions of the experiments whenever possible. No animals or data points were excluded from the analysis. Normalcy was assessed using Shapiro–Wilk test. Equal variances were assessed using F test or Bartlett’s test. Statistical significance was tested using two-tailed unpaired t-test, Welch’s t-test, one-way analysis of variance (ANOVA), and two-way ANOVA followed by Tukey’s or Bonferroni post hoc test, wherever appropriate. Data are presented as mean ± standard error of the mean. p < 0.05 was considered significant. Significance is marked as *p < 0.05, **p < 0.01, and ***p <0.001.
Data availability
All data generated or analyzed during this study are included in the manuscript. Source data have been provided for Figures 1 and 2.
References
-
Cortical excitatory neurons and glia, but not GABAergic neurons, are produced in the Emx1-expressing lineageThe Journal of Neuroscience 22:6309–6314.https://doi.org/10.1523/JNEUROSCI.22-15-06309.2002
-
Immunological markers for central nervous system gliaNeuroscience Bulletin 39:379–392.https://doi.org/10.1007/s12264-022-00938-2
-
Identification of Tmem10 as a novel late-stage oligodendrocytes marker for detecting hypomyelinationInternational Journal of Biological Sciences 10:33–42.https://doi.org/10.7150/ijbs.7526
-
Dorsally and ventrally derived oligodendrocytes have similar electrical properties but myelinate preferred tractsThe Journal of Neuroscience 31:6809–6819.https://doi.org/10.1523/JNEUROSCI.6474-10.2011
-
Fate mapping Nkx2.1-lineage cells in the mouse telencephalonThe Journal of Comparative Neurology 506:16–29.https://doi.org/10.1002/cne.21529
-
Opalin, a transmembrane sialylglycoprotein located in the central nervous system myelin paranodal loop membraneThe Journal of Biological Chemistry 283:20830–20840.https://doi.org/10.1074/jbc.M801314200
Article and author information
Author details
Funding
National Science and Technology Innovation 2030 Major Projects of China (STI2030-Major Projects-2022ZD0206500)
- Miao He
National Natural Science Foundation of China (32171087)
- Miao He
National Natural Science Foundation of China (32371145)
- Ling Gong
The funders had no role in study design, data collection, and interpretation, or the decision to submit the work for publication.
Acknowledgements
We thank Dr. Yilin Tai for helpful discussion and Dr. Min Jiang from core facility of IOBS, Fudan University, for technical support on imaging experiments. This study was supported by funds from the National Science and Technology Innovation 2030 Major Projects of China (STI2030-Major Projects-2022ZD0206500), National Natural Science Foundation of China (32171087, 32371145).
Ethics
All mouse studies were carried out in strict accordance with the guidelines of the Institutional Animal Care and Use Committee of School of Basic Medical Sciences, Fudan University. All husbandry and experimental procedures were reviewed and approved by the same committee (Permit Number: 20210302-137). All applicable institutional and/or national guidelines for the care and use of animals were followed.
Version history
- Sent for peer review:
- Preprint posted:
- Reviewed Preprint version 1:
- Reviewed Preprint version 2:
- Version of Record published:
Cite all versions
You can cite all versions using the DOI https://doi.org/10.7554/eLife.95406. This DOI represents all versions, and will always resolve to the latest one.
Copyright
© 2024, Cai, Zhao, Shi et al.
This article is distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use and redistribution provided that the original author and source are credited.
Metrics
-
- 1,127
- views
-
- 131
- downloads
-
- 2
- citations
Views, downloads and citations are aggregated across all versions of this paper published by eLife.
Download links
A two-part list of links to download the article, or parts of the article, in various formats.
Downloads (link to download the article as PDF)
Open citations (links to open the citations from this article in various online reference manager services)
Cite this article (links to download the citations from this article in formats compatible with various reference manager tools)
Embryonic origins of forebrain oligodendrocytes revisited by combinatorial genetic fate mapping
eLife 13:RP95406.
https://doi.org/10.7554/eLife.95406.3
Further reading
-
- Developmental Biology
Wing dimorphism is a common phenomenon that plays key roles in the environmental adaptation of aphid; however, the signal transduction in response to environmental cues and the regulation mechanism related to this event remain unknown. Adenosine (A) to inosine (I) RNA editing is a post-transcriptional modification that extends transcriptome variety without altering the genome, playing essential roles in numerous biological and physiological processes. Here, we present a chromosome-level genome assembly of the rose-grain aphid Metopolophium dirhodum by using PacBio long HiFi reads and Hi-C technology. The final genome assembly for M. dirhodum is 447.8 Mb, with 98.50% of the assembled sequences anchored to nine chromosomes. The contig and scaffold N50 values are 7.82 and 37.54 Mb, respectively. A total of 18,003 protein-coding genes were predicted, of which 92.05% were functionally annotated. In addition, 11,678 A-to-I RNA-editing sites were systematically identified based on this assembled M. dirhodum genome, and two synonymous A-to-I RNA-editing sites on CYP18A1 were closely associated with transgenerational wing dimorphism induced by crowding. One of these A-to-I RNA-editing sites may prevent the binding of miR-3036-5p to CYP18A1, thus elevating CYP18A1 expression, decreasing 20E titer, and finally regulating the wing dimorphism of offspring. Meanwhile, crowding can also inhibit miR-3036-5p expression and further increase CYP18A1 abundance, resulting in winged offspring. These findings support that A-to-I RNA editing is a dynamic mechanism in the regulation of transgenerational wing dimorphism in aphids and would advance our understanding of the roles of RNA editing in environmental adaptability and phenotypic plasticity.
-
- Developmental Biology
The evolutionarily conserved Hippo (Hpo) pathway has been shown to impact early development and tumorigenesis by governing cell proliferation and apoptosis. However, its post-developmental roles are relatively unexplored. Here, we demonstrate its roles in post-mitotic cells by showing that defective Hpo signaling accelerates age-associated structural and functional decline of neurons in Caenorhabditis elegans. Loss of wts-1/LATS, the core kinase of the Hpo pathway, resulted in premature deformation of touch neurons and impaired touch responses in a yap-1/YAP-dependent manner, the downstream transcriptional co-activator of LATS. Decreased movement as well as microtubule destabilization by treatment with colchicine or disruption of microtubule-stabilizing genes alleviated the neuronal deformation of wts-1 mutants. Colchicine exerted neuroprotective effects even during normal aging. In addition, the deficiency of a microtubule-severing enzyme spas-1 also led to precocious structural deformation. These results consistently suggest that hyper-stabilized microtubules in both wts-1-deficient neurons and normally aged neurons are detrimental to the maintenance of neuronal structural integrity. In summary, Hpo pathway governs the structural and functional maintenance of differentiated neurons by modulating microtubule stability, raising the possibility that the microtubule stability of fully developed neurons could be a promising target to delay neuronal aging. Our study provides potential therapeutic approaches to combat age- or disease-related neurodegeneration.




{kind=link}
{kind=link}
{kind=link}